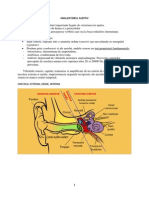
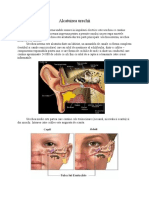
S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Sunetul fizicii: Acustica fenomenologicăDe la EverandSunetul fizicii: Acustica fenomenologicăEvaluare: 4.5 din 5 stele4.5/5 (2)
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Rezidentiat Vol 2Document381 paginiRezidentiat Vol 2Madalina Luca100% (2)
- D Antonescu Elemente de Ortopedie Si TraumatologieDocument422 paginiD Antonescu Elemente de Ortopedie Si TraumatologieIulia Niculae100% (8)
- Carte CURS MEDICINA DE FAMILIE DR Celina Stafie (26 Aprilie 2016) PDFDocument236 paginiCarte CURS MEDICINA DE FAMILIE DR Celina Stafie (26 Aprilie 2016) PDFIoana Avram100% (3)
- Receptia AuditivaDocument10 paginiReceptia AuditivaRadu BulboacaÎncă nu există evaluări
- Defectologie Si LogopedieDocument77 paginiDefectologie Si LogopedieAnca Maria MihuÎncă nu există evaluări
- UrecheaDocument6 paginiUrecheaTeodor Mihai100% (2)
- Biofizica Receptiei AuditiveDocument6 paginiBiofizica Receptiei AuditiveSharak NestÎncă nu există evaluări
- Sunetul Si Perceptia AcestuiaDocument18 paginiSunetul Si Perceptia AcestuiatestusertestÎncă nu există evaluări
- Otita 2021Document55 paginiOtita 2021Andreea Ciubotaru100% (2)
- Cursuri ORL AMGDocument78 paginiCursuri ORL AMGAndaOprea67% (6)
- Mecanismul AuzuluiDocument4 paginiMecanismul AuzuluiHandMadeÎncă nu există evaluări
- BIOACUSTICADocument8 paginiBIOACUSTICAKasai Maria Petronela100% (3)
- Fiziologia Sistemului AuditivDocument20 paginiFiziologia Sistemului AuditivOana Cristina100% (1)
- M1 Anatomia Și Fiziologia Omului - Fiziologia Analizatorului Auditiv Și VestibularDocument7 paginiM1 Anatomia Și Fiziologia Omului - Fiziologia Analizatorului Auditiv Și VestibularFlorina Alina LicaÎncă nu există evaluări
- Anatomia Si Fiziologia Analizatorului AuditivDocument7 paginiAnatomia Si Fiziologia Analizatorului AuditivAnabela Alexandra100% (1)
- Analizatorul AuditivDocument18 paginiAnalizatorul AuditiveugenalexandrovÎncă nu există evaluări
- Urechea. Aparat AuditivDocument4 paginiUrechea. Aparat AuditivRomanescu George100% (1)
- Curs Asist 2 Final BT PDFDocument200 paginiCurs Asist 2 Final BT PDFIonutz VargaÎncă nu există evaluări
- Anatomia UrechiiDocument16 paginiAnatomia UrechiiChGabriellaÎncă nu există evaluări
- Analizatorul Auditiv Este Organul de Simt Care Asigura Perceperea Unde Lor SonoreDocument3 paginiAnalizatorul Auditiv Este Organul de Simt Care Asigura Perceperea Unde Lor SonoreAndreea BchsÎncă nu există evaluări
- Anatomia, Fiziologia, Patologia Organelor de Auz, Văz Și Vorbire PDFDocument74 paginiAnatomia, Fiziologia, Patologia Organelor de Auz, Văz Și Vorbire PDFanushka1988Încă nu există evaluări
- Proiect Urechea BionicaDocument7 paginiProiect Urechea BionicaFlorin PîrîialăÎncă nu există evaluări
- Subiecte Rezolvate OMF IasiDocument20 paginiSubiecte Rezolvate OMF IasiAndreea VelnicÎncă nu există evaluări
- Valori Normale Ale Unor Indici in PediatrieDocument16 paginiValori Normale Ale Unor Indici in PediatrieOlga Sazonova-GherasimÎncă nu există evaluări
- Neurobiologia Auditiei - Laborator 1Document10 paginiNeurobiologia Auditiei - Laborator 1idaminu2321Încă nu există evaluări
- Biofizica Receptiei AuditiveDocument14 paginiBiofizica Receptiei AuditiveAndrone Camelia100% (1)
- Orl Si Nursing in Orl Suport de Curs AnuDocument32 paginiOrl Si Nursing in Orl Suport de Curs Anumirela rusuÎncă nu există evaluări
- Cursuri ORL - OdtDocument78 paginiCursuri ORL - OdtAndaOpreaÎncă nu există evaluări
- Lumea SunetelorDocument11 paginiLumea SunetelorCristina Maria DeneşanÎncă nu există evaluări
- Notiuni de Anatomie Si Fizionomie A UrechiiDocument15 paginiNotiuni de Anatomie Si Fizionomie A UrechiiCrina Maria PuiaÎncă nu există evaluări
- Biofizica Recepţiei AuditiveDocument22 paginiBiofizica Recepţiei AuditiveValentino PetreÎncă nu există evaluări
- Analizatorul AuditivDocument14 paginiAnalizatorul Auditivmiki.elaÎncă nu există evaluări
- Analizatorul AuditivDocument7 paginiAnalizatorul AuditivSmadu AdrianaÎncă nu există evaluări
- LihjDocument74 paginiLihjanushka1988Încă nu există evaluări
- Analizatorul AuditivDocument15 paginiAnalizatorul Auditivcrr_29Încă nu există evaluări
- 8.fiziologia Aparatului de Transmitere A SunetelorDocument3 pagini8.fiziologia Aparatului de Transmitere A SunetelorAndrei RotaruÎncă nu există evaluări
- 1 AuditivDocument2 pagini1 AuditivRussu DumitruÎncă nu există evaluări
- URECHEADocument25 paginiURECHEAToma GeorgianaaÎncă nu există evaluări
- UrecheaDocument22 paginiUrechearaysa_svÎncă nu există evaluări
- Analizatorul AuditivDocument5 paginiAnalizatorul AuditivCristina LoveÎncă nu există evaluări
- Biofizica Receptiei AuditiveDocument18 paginiBiofizica Receptiei AuditiveIoana Mihaela SerbanÎncă nu există evaluări
- Organul AuzuluiDocument21 paginiOrganul AuzuluiLilia LapteacruÎncă nu există evaluări
- Rezumat La Ureche Pentru BiologieDocument3 paginiRezumat La Ureche Pentru BiologieAdrian BejenaruÎncă nu există evaluări
- Analizatorul AuditivDocument2 paginiAnalizatorul AuditivSilviuu VîjialaÎncă nu există evaluări
- 1391Document76 pagini1391Silviu100% (1)
- Biofizica Recepţiei AuditiveDocument19 paginiBiofizica Recepţiei AuditiveMihai ArcaleanÎncă nu există evaluări
- UrecheaDocument3 paginiUrecheaCamelia CammyÎncă nu există evaluări
- Aparatul Auditiv Si VizualDocument8 paginiAparatul Auditiv Si VizualAndra Apostol100% (1)
- Urechea UmanaDocument2 paginiUrechea UmanaMolnarRaduÎncă nu există evaluări
- Cum AuzimDocument19 paginiCum AuzimAlexandru ClisuÎncă nu există evaluări
- Biomecanica Sistemului AuditivDocument8 paginiBiomecanica Sistemului AuditivStefania FrincuÎncă nu există evaluări
- Titlul Lucrarii - Organul de Simt - UrecheaDocument5 paginiTitlul Lucrarii - Organul de Simt - UrecheaDaniel NicolaeÎncă nu există evaluări
- WWW - Referat.ro-Aparatul Auditiv - Doc63da1 IONELADocument2 paginiWWW - Referat.ro-Aparatul Auditiv - Doc63da1 IONELADinu ValentinÎncă nu există evaluări
- Auditiv Și VestibularDocument8 paginiAuditiv Și VestibularLarisa DumitruÎncă nu există evaluări
- Urechea. Aparat AuditivDocument2 paginiUrechea. Aparat AuditivElena RașcovÎncă nu există evaluări
- Urechea Echilibru ProiectDocument17 paginiUrechea Echilibru ProiectCrina MariaÎncă nu există evaluări
- Urechea UmanaDocument8 paginiUrechea UmanaAnd DelÎncă nu există evaluări
- VIII - AuditivDocument21 paginiVIII - AuditivDiana ZețuÎncă nu există evaluări
- UrecheaDocument2 paginiUrecheaMihai MihaiÎncă nu există evaluări
- Urechea Umană Și Sunetul În FizicăDocument11 paginiUrechea Umană Și Sunetul În FizicăAlex OpreaÎncă nu există evaluări
- Vaccinuri de Necesitate EpidemiologicaDocument38 paginiVaccinuri de Necesitate EpidemiologicaScolobiuc Emma ElenaÎncă nu există evaluări
- Imobilizarea Leziunilor OsteoarticulareDocument8 paginiImobilizarea Leziunilor OsteoarticulareScolobiuc Emma ElenaÎncă nu există evaluări
- Pro 4820 20.07.12Document6 paginiPro 4820 20.07.12Caciula AurasÎncă nu există evaluări
- Curriculum Rezidentiat Chirurgie GeneralaDocument18 paginiCurriculum Rezidentiat Chirurgie Generalatynaionescu7124Încă nu există evaluări
- SL6 Chirurgie CardiovascularaDocument1 paginăSL6 Chirurgie CardiovascularaScolobiuc Emma ElenaÎncă nu există evaluări
- Psihiatrie SubiecteDocument12 paginiPsihiatrie SubiecteOrtieÎncă nu există evaluări
- Microbi StagiuDocument77 paginiMicrobi StagiuScolobiuc Emma ElenaÎncă nu există evaluări
- RinaDocument4 paginiRinaLauraFlorescuÎncă nu există evaluări
- Lista Cursuri Si Prelegeri Pana in S12Document1 paginăLista Cursuri Si Prelegeri Pana in S12Scolobiuc Emma ElenaÎncă nu există evaluări
- Intox. AcuteDocument134 paginiIntox. AcuteScolobiuc Emma ElenaÎncă nu există evaluări
- Formule SPMDocument1 paginăFormule SPMDiana IacobÎncă nu există evaluări
- Toxicologie GeneralaDocument51 paginiToxicologie GeneralaAndreea GheorgheÎncă nu există evaluări
- A Petris Note de Stagiu Ed Pim Iasi 2008 Barem ECGDocument41 paginiA Petris Note de Stagiu Ed Pim Iasi 2008 Barem ECGMerry LeeÎncă nu există evaluări
- Cur SuriDocument1 paginăCur SuriScolobiuc Emma ElenaÎncă nu există evaluări
- Astmul BronsicDocument86 paginiAstmul BronsiccristianÎncă nu există evaluări
- Grile Licenţă - Medicină - Limba RomânăDocument168 paginiGrile Licenţă - Medicină - Limba Românăandreea100% (1)
- Infarctul Miocardic Acut 2515968117126518Document135 paginiInfarctul Miocardic Acut 2515968117126518Oana Cristina VoineaÎncă nu există evaluări
- CitateDocument1 paginăCitateScolobiuc Emma ElenaÎncă nu există evaluări
- Curs 26Document3 paginiCurs 26Simona CiobanasuÎncă nu există evaluări
- Consultatia PrenatalaDocument5 paginiConsultatia PrenatalaScolobiuc Emma ElenaÎncă nu există evaluări
- EpidemioDocument6 paginiEpidemioScolobiuc Emma ElenaÎncă nu există evaluări
- Studiu Privind Frecventa Intoxicatiilor Acute La CopiiDocument7 paginiStudiu Privind Frecventa Intoxicatiilor Acute La CopiiScolobiuc Emma ElenaÎncă nu există evaluări
- Curs 1Document1 paginăCurs 1Elisa StanÎncă nu există evaluări
- Curs 2. Streptococ Meningococ 2013 2014Document33 paginiCurs 2. Streptococ Meningococ 2013 2014Ciopraga IuliaÎncă nu există evaluări
- Tematica Geriatrie AN VIDocument1 paginăTematica Geriatrie AN VIScolobiuc Emma ElenaÎncă nu există evaluări