S-ar putea să vă placă și
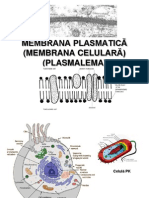
- Membrana PlasmaticăDocument56 paginiMembrana PlasmaticăAndrada Raicu100% (1)
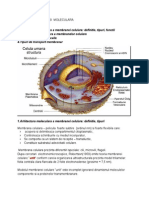
- Membrana CelularaDocument25 paginiMembrana CelularaFlavius DobreÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- Elemente de Transport Prin Membrana CelularaDocument29 paginiElemente de Transport Prin Membrana CelularaIoana CojocaruÎncă nu există evaluări
- BIOCEL Sinteza-1-1Document45 paginiBIOCEL Sinteza-1-1Adelin FrumusheluÎncă nu există evaluări
- Celula EucariotaDocument15 paginiCelula EucariotaveronicaÎncă nu există evaluări
- Membrana CelularaDocument9 paginiMembrana CelularaRadu MaziluÎncă nu există evaluări
- Membrana Celulara PDFDocument40 paginiMembrana Celulara PDFOzana CopyShop100% (1)
- Sistemul Nervos - Actul Şi Arcul Reflex 2Document55 paginiSistemul Nervos - Actul Şi Arcul Reflex 2Alexandru ChivaranÎncă nu există evaluări
- 1.fiziologia Celulei VegetaleDocument6 pagini1.fiziologia Celulei VegetaleDragan GabrielaÎncă nu există evaluări
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 paginiATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- C04 Membrana Celulara JonctiuniDocument46 paginiC04 Membrana Celulara JonctiunipanterabloÎncă nu există evaluări
- Perioada Pre-EmbrionaraDocument2 paginiPerioada Pre-EmbrionaraCristian MariusÎncă nu există evaluări
- Difuzia SimplaDocument1 paginăDifuzia SimplaTeodora Elena100% (1)
- PinocitozaDocument26 paginiPinocitozaMonicaÎncă nu există evaluări
- 1b Lanțul Transportor de ElectroniDocument82 pagini1b Lanțul Transportor de ElectroniLaura HutzelÎncă nu există evaluări
- Endocitoza Mediată de ReceptoriDocument13 paginiEndocitoza Mediată de ReceptoriBianca BibanÎncă nu există evaluări
- Functiile SangeluiDocument3 paginiFunctiile SangeluiEduard ButuÎncă nu există evaluări
- Diferentiere CelularaDocument30 paginiDiferentiere CelularaMihaela Anton100% (1)
- Embriologie Curs 5 Saptamana 3Document71 paginiEmbriologie Curs 5 Saptamana 3LumyÎncă nu există evaluări
- C 6, 7 - Enzime (Slide 15)Document38 paginiC 6, 7 - Enzime (Slide 15)Elena2130Încă nu există evaluări
- Citoscheletul PDFDocument41 paginiCitoscheletul PDFOzana CopyShopÎncă nu există evaluări
- Cilii Si FlagelulDocument1 paginăCilii Si FlagelulBarbie AndreeaÎncă nu există evaluări
- Celula Anatomie MedicinaDocument27 paginiCelula Anatomie MedicinaManu ElaÎncă nu există evaluări
- CitoscheletulDocument6 paginiCitoscheletulJessica Smith100% (1)
- Enzime 1Document38 paginiEnzime 1Elena Alina TÎncă nu există evaluări
- Lizozomi Curs TextDocument10 paginiLizozomi Curs TextFlorin DumitracheÎncă nu există evaluări
- Curs 3 Aparatul GolgiDocument4 paginiCurs 3 Aparatul GolgiOana Koppandi100% (1)
- Schimbul de Substanțe Dintre Celulă Și Mediul AmbiantDocument5 paginiSchimbul de Substanțe Dintre Celulă Și Mediul AmbiantRadu MarianÎncă nu există evaluări
- Diviziunea CelularaDocument29 paginiDiviziunea CelularaCrinablue100% (1)
- WWW - Referat.ro Plastidele 3f7ebDocument8 paginiWWW - Referat.ro Plastidele 3f7ebErik MartinezÎncă nu există evaluări
- GAMETOGENEZA Spermatogeneza OvogenezaDocument10 paginiGAMETOGENEZA Spermatogeneza OvogenezaAntonio SpițaÎncă nu există evaluări
- BiocelDocument24 paginiBiocelficioruÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Citologie AnimalaDocument28 paginiCitologie AnimalaEmilia Sabau100% (1)
- 6 HematopoezaDocument8 pagini6 HematopoezaValentin MoisăÎncă nu există evaluări
- Morfologia Plantelor - Botanica Semestrul 1Document12 paginiMorfologia Plantelor - Botanica Semestrul 1David StoianÎncă nu există evaluări
- Plasma SanguinaDocument6 paginiPlasma SanguinadenisemichelaÎncă nu există evaluări
- Nucleolul Şi Funcţiile LuiDocument2 paginiNucleolul Şi Funcţiile LuiNutza Zadic100% (1)
- Celula Diviziunea CelularaDocument26 paginiCelula Diviziunea CelularaGabØrÎncă nu există evaluări
- 3 Studiul Celulei - Forma Si Dimensiunile CeluleiDocument3 pagini3 Studiul Celulei - Forma Si Dimensiunile CeluleiAndrEea MIronÎncă nu există evaluări
- Structura ProteinelorDocument13 paginiStructura ProteinelorA B100% (1)
- Rolul Reticulului Endoplasmatic În DetoxifiereDocument15 paginiRolul Reticulului Endoplasmatic În DetoxifiereAdelina MkyÎncă nu există evaluări
- Matrice ExtracelularaDocument27 paginiMatrice ExtracelularaOana PatriciaÎncă nu există evaluări
- Citologie MembranaDocument33 paginiCitologie MembranaOsman Daniela100% (1)
- Caracterizarea ProteinelorDocument3 paginiCaracterizarea ProteinelorRaluca Sofian100% (1)
- SangeleDocument6 paginiSangelelidiacampeanÎncă nu există evaluări
- Curs 2 Clasificare. HemoglobinaDocument20 paginiCurs 2 Clasificare. Hemoglobinajacksparrow2Încă nu există evaluări
- TAM4Document37 paginiTAM4Adriana Loredana SecrieriuÎncă nu există evaluări
- 4 Transport Membranar MGDocument19 pagini4 Transport Membranar MGmarkopavloski100% (1)
- BiocelDocument26 paginiBiocelAdelin FrumusheluÎncă nu există evaluări
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- Membrana Celulara Structura Moleculara Si Functiile Membranelor CelulareDocument25 paginiMembrana Celulara Structura Moleculara Si Functiile Membranelor CelulareAna CirsteaÎncă nu există evaluări
- Membrane Biologice - BunDocument21 paginiMembrane Biologice - BunMarius Alexa67% (3)
- Subiecte Biocel 2015Document43 paginiSubiecte Biocel 2015Daniel Mihai GalbenusÎncă nu există evaluări
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Membrane Biologice 1Document10 paginiMembrane Biologice 1Anca NegreanÎncă nu există evaluări
- Catb 5Document3 paginiCatb 5Diana ValentinaÎncă nu există evaluări
- Curs 4 BIOFIZDocument72 paginiCurs 4 BIOFIZfloricapopovici168Încă nu există evaluări
- Modelul MLM de BusinessDocument2 paginiModelul MLM de BusinessClaudiu BudÎncă nu există evaluări
- Boris Cyrulnik - Rușinea. Mai Bine Mor Decât Să Spun PDFDocument241 paginiBoris Cyrulnik - Rușinea. Mai Bine Mor Decât Să Spun PDFamad4you100% (1)
- AntiacideDocument1 paginăAntiacideClaudiu BudÎncă nu există evaluări
- Grile Examen FarmacologieDocument3 paginiGrile Examen FarmacologieClaudiu Bud100% (1)
- Stefan Cel MareDocument2 paginiStefan Cel MareClaudiu BudÎncă nu există evaluări
- MITOCONDRIADocument5 paginiMITOCONDRIAClaudiu Bud100% (1)
- Fiz AleDocument7 paginiFiz AleClaudiu BudÎncă nu există evaluări
- Secretele Mintii de Milionar T HARV EKER NetDocument23 paginiSecretele Mintii de Milionar T HARV EKER NetMarya YoanaÎncă nu există evaluări
- Nivelele Unei Limbi StraineDocument4 paginiNivelele Unei Limbi StraineClaudiu BudÎncă nu există evaluări
- Noțiuni Elementare de ContabilitateDocument19 paginiNoțiuni Elementare de ContabilitateClaudiu BudÎncă nu există evaluări
- Transportul Transmembranar Al Macromoleculelor. Exocitoza Şi EndocitozaDocument6 paginiTransportul Transmembranar Al Macromoleculelor. Exocitoza Şi EndocitozaClaudiu BudÎncă nu există evaluări
- ReticulDocument4 paginiReticulClaudiu BudÎncă nu există evaluări
- Hematologie Farmacie Curs AnemiiDocument97 paginiHematologie Farmacie Curs AnemiiClaudiu BudÎncă nu există evaluări
- Referat Info ClauDocument4 paginiReferat Info ClauClaudiu BudÎncă nu există evaluări
- Maria Treben - Sanatate Din Farmacia Domnului (A4, Doc - 134pag, PDF - 161 Pag Pt. Ca Scribd Converteste Rau in PDF, Digital, Concordanta Pagini)Document166 paginiMaria Treben - Sanatate Din Farmacia Domnului (A4, Doc - 134pag, PDF - 161 Pag Pt. Ca Scribd Converteste Rau in PDF, Digital, Concordanta Pagini)dlucian82100% (3)
- New Microsoft Office Word DocumentDocument9 paginiNew Microsoft Office Word DocumentClaudiu BudÎncă nu există evaluări
- Anatomia Aparatului RespiratorDocument15 paginiAnatomia Aparatului RespiratorClaudiu BudÎncă nu există evaluări
- Anatomia Aparatului RespiratorDocument15 paginiAnatomia Aparatului RespiratorClaudiu BudÎncă nu există evaluări
- Cheile Magice Ale Lui SolomonDocument17 paginiCheile Magice Ale Lui SolomonBelieveit Ro70% (10)
- Aerosol IDocument2 paginiAerosol IClaudiu BudÎncă nu există evaluări
- Termodinamica - Referat MediocruDocument4 paginiTermodinamica - Referat MediocruClaudiu Bud100% (1)
- Cheile Magice Ale Lui SolomonDocument17 paginiCheile Magice Ale Lui SolomonBelieveit Ro70% (10)
- Referat Mocan Mihai AndreiDocument11 paginiReferat Mocan Mihai AndreiClaudiu BudÎncă nu există evaluări
- Citate ValeDocument1 paginăCitate ValeClaudiu BudÎncă nu există evaluări