S-ar putea să vă placă și
- Dobre Amelia KINETOTERAPIA DEFICIENTELOR FIZICE SI SENZORIALEDocument4 paginiDobre Amelia KINETOTERAPIA DEFICIENTELOR FIZICE SI SENZORIALEAmelia F. DobreÎncă nu există evaluări
- Ioana Pearsica - Relatia Actor-Spectator La GrotowskiDocument8 paginiIoana Pearsica - Relatia Actor-Spectator La GrotowskiAntonia IordacheÎncă nu există evaluări
- C 1 Sistemul NeuromuscularDocument22 paginiC 1 Sistemul NeuromuscularDaniela Acceleanu100% (1)
- DVMDocument30 paginiDVMValentin Bogdan VfbÎncă nu există evaluări
- Chirped LessDocument158 paginiChirped LessLemmyÎncă nu există evaluări
- Sindroamele Si Afectiunile ExtrapiramidaleDocument39 paginiSindroamele Si Afectiunile ExtrapiramidaleanaebritÎncă nu există evaluări
- Exercitiul Fizic În Profilaxia Si Tratamentul Procesului de ImbatranireDocument6 paginiExercitiul Fizic În Profilaxia Si Tratamentul Procesului de ImbatranireVas Raul-MihaiÎncă nu există evaluări
- SUBIECTE Recuperare NeurologieDocument3 paginiSUBIECTE Recuperare NeurologieTurta AdyÎncă nu există evaluări
- AruncariDocument38 paginiAruncaricriminalistÎncă nu există evaluări
- DefinitieDocument1 paginăDefinitieMaria AdÎncă nu există evaluări
- NociceptiaDocument17 paginiNociceptiaVica LazariÎncă nu există evaluări
- Leziuni Musculare de SuprasolicitareDocument16 paginiLeziuni Musculare de SuprasolicitareValentin Bogdan VfbÎncă nu există evaluări
- Fortele Interioare Ale LocomotieiDocument8 paginiFortele Interioare Ale LocomotieiStefan ToaderÎncă nu există evaluări
- Miologie CursDocument18 paginiMiologie Cursblizzard90Încă nu există evaluări
- Referat Mortean VladimirDocument17 paginiReferat Mortean VladimirVladimir MorteanÎncă nu există evaluări
- Anatomie Si Biomecanica UmarDocument28 paginiAnatomie Si Biomecanica UmarHristodoulakis Stefanos0% (1)
- KinesiologieDocument7 paginiKinesiologieValeria StillÎncă nu există evaluări
- Mușchi Origine Insertie Actiune InervatieDocument6 paginiMușchi Origine Insertie Actiune InervatieFlorinel OpreaÎncă nu există evaluări
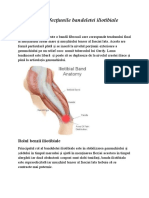
- Afectiunile BandeleteiDocument16 paginiAfectiunile BandeleteiAlexandra CroitoruÎncă nu există evaluări
- Refacerea Dupa Efort ArticolDocument2 paginiRefacerea Dupa Efort Articolkarl_bluewishÎncă nu există evaluări
- Fiziopatologia AntrenamentuluiDocument11 paginiFiziopatologia AntrenamentuluiGeanina ButunoiÎncă nu există evaluări
- RECEPTORIIDocument3 paginiRECEPTORIIAlexe DianaÎncă nu există evaluări
- Asistenta Psihologica in KinetoterapieDocument4 paginiAsistenta Psihologica in KinetoterapieFlorin DinuÎncă nu există evaluări
- Clavicula Si OmoplatulDocument10 paginiClavicula Si OmoplatulAlina CamerzanÎncă nu există evaluări
- Contractia Musculaturii Muschiului CardiacDocument4 paginiContractia Musculaturii Muschiului CardiacMitrache AureliaÎncă nu există evaluări
- Sistem MuschiDocument19 paginiSistem MuschiDan MassonÎncă nu există evaluări
- Neuro BunDocument41 paginiNeuro Bunpunktuletzz100% (1)
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- FiziopatologiaDocument9 paginiFiziopatologiachivumihaelaÎncă nu există evaluări
- KinetologieDocument15 paginiKinetologiejulie77eÎncă nu există evaluări
- Luxatia Congenitala de SoldDocument11 paginiLuxatia Congenitala de SoldPintilie AlexandraÎncă nu există evaluări
- Dorina Flora Tehnici de Baza in Kinetoterapie PDFDocument327 paginiDorina Flora Tehnici de Baza in Kinetoterapie PDFIsabela IacobÎncă nu există evaluări
- Muschiul StriatDocument7 paginiMuschiul StriatDima SilviuÎncă nu există evaluări
- Profesia de KinetoterapeutDocument2 paginiProfesia de KinetoterapeutMihai AcăeiÎncă nu există evaluări
- Subiecte KinesiologieDocument6 paginiSubiecte KinesiologieDaniel Nitescu100% (1)
- Caile Sensibilitatii La Cap Si La Gat CURS 4Document4 paginiCaile Sensibilitatii La Cap Si La Gat CURS 4Alin PacurarÎncă nu există evaluări
- Coada de CalDocument4 paginiCoada de CalAdi RughiÎncă nu există evaluări
- RELAXAREADocument34 paginiRELAXAREAAdrian VladÎncă nu există evaluări
- AlpinismulDocument56 paginiAlpinismulElena TonuÎncă nu există evaluări
- 8 Circuite Neuronale 2Document1 pagină8 Circuite Neuronale 2Alex Cu TridentÎncă nu există evaluări
- Traumatismele Maduvei SpinariiDocument27 paginiTraumatismele Maduvei SpinariiIonut Si Madalina PatrascuÎncă nu există evaluări
- Functia de Conducere A Trunchiului CerebralDocument19 paginiFunctia de Conducere A Trunchiului Cerebral12345Încă nu există evaluări
- Curs 3 - Generalitati Despre Muschi PDFDocument60 paginiCurs 3 - Generalitati Despre Muschi PDFFlorinela PopaÎncă nu există evaluări
- HemodinamicaDocument28 paginiHemodinamicazxcvnbvnÎncă nu există evaluări
- Sindromul ExtrapiramidalDocument27 paginiSindromul ExtrapiramidalFlorentina Kociopolos CiobanuÎncă nu există evaluări
- Universitatea Spiru Haret Olga DjamoDocument30 paginiUniversitatea Spiru Haret Olga Djamocosmin_mirelÎncă nu există evaluări
- Educatie Fizica Sport Si KinetoterapieDocument200 paginiEducatie Fizica Sport Si KinetoterapieMironescu LaviniaÎncă nu există evaluări
- Fiziologie TotDocument34 paginiFiziologie TotbamsweÎncă nu există evaluări
- TMK Curs ViiiDocument10 paginiTMK Curs ViiiviorelÎncă nu există evaluări
- Curs 5Document14 paginiCurs 5sham harbaÎncă nu există evaluări
- FiziologieDocument47 paginiFiziologieGeorgian SerafimÎncă nu există evaluări
- Anatomia Si Fiziologia Sistemului MuscularDocument36 paginiAnatomia Si Fiziologia Sistemului MuscularGligan MihaiÎncă nu există evaluări
- Îfiziologia Sist MuscDocument11 paginiÎfiziologia Sist MuscmalexandraÎncă nu există evaluări
- Tema 1 BGKDocument10 paginiTema 1 BGKDorian T FlorinÎncă nu există evaluări
- MuscularDocument46 paginiMuscularCatalin CusnirÎncă nu există evaluări
- Fiziologia MuschilorDocument24 paginiFiziologia Muschilorlena red100% (1)
- CURS MUSCHI FIZIO PPTDocument46 paginiCURS MUSCHI FIZIO PPTIrina MureșanÎncă nu există evaluări
- Sistemului NeuromuscularDocument54 paginiSistemului NeuromuscularStefi MaryÎncă nu există evaluări
- Referat Anatomie 3Document6 paginiReferat Anatomie 3Mihnea BucurÎncă nu există evaluări
- Sistemul MuscularDocument9 paginiSistemul MuscularDiana AndreeaÎncă nu există evaluări
- Institutii Ale Uniunii EuropeneDocument11 paginiInstitutii Ale Uniunii EuropeneEu VictorÎncă nu există evaluări
- Anexa 1 - Asamblari DemontabileDocument1 paginăAnexa 1 - Asamblari DemontabileEu VictorÎncă nu există evaluări
- Curs 13 - 13 LPDocument20 paginiCurs 13 - 13 LPSimionHessÎncă nu există evaluări
- Coroziunea GalvanicaDocument2 paginiCoroziunea GalvanicaCatalin DumitruÎncă nu există evaluări
- AnoziDocument3 paginiAnoziFlorin MuşatÎncă nu există evaluări
- Peda GogiePeda GogiePeda GogiePeda GogiePeda GogiePeda GogieDocument2 paginiPeda GogiePeda GogiePeda GogiePeda GogiePeda GogiePeda GogieEu VictorÎncă nu există evaluări
- Principiul Minimului de Energie Si Maximul de EntropieDocument9 paginiPrincipiul Minimului de Energie Si Maximul de EntropieCristina MoldoveanuÎncă nu există evaluări
- Conductia Extrinseca A SemiconductorilorDocument1 paginăConductia Extrinseca A SemiconductorilorEu VictorÎncă nu există evaluări
- Bioingineria CDocument1 paginăBioingineria CEu VictorÎncă nu există evaluări
- Copiute MEEDocument3 paginiCopiute MEEEu VictorÎncă nu există evaluări
- Lista Lucrarilor Practice de LaboratorDocument1 paginăLista Lucrarilor Practice de LaboratorEu VictorÎncă nu există evaluări
- SUBIECTE Chimie 2011 ELTHDocument11 paginiSUBIECTE Chimie 2011 ELTHRaduCiciuÎncă nu există evaluări
- 3 Calculul Parametrilor InfasurariiDocument13 pagini3 Calculul Parametrilor InfasurariiMihai ConstantinescuÎncă nu există evaluări
- Proiect MP 93Document6 paginiProiect MP 93Eu VictorÎncă nu există evaluări
- Chimie Grafic Duritatea ApeiDocument2 paginiChimie Grafic Duritatea ApeiEu VictorÎncă nu există evaluări
- Caracteristici ExterneDocument3 paginiCaracteristici ExterneEu VictorÎncă nu există evaluări
- T1 - Transformator Monofazat PDFDocument2 paginiT1 - Transformator Monofazat PDFNicu IlieÎncă nu există evaluări
- Proiect MPDocument14 paginiProiect MPEu VictorÎncă nu există evaluări
- Radarul IstoricDocument2 paginiRadarul IstoricEu VictorÎncă nu există evaluări
- Probleme TDEE 2015Document8 paginiProbleme TDEE 2015Eu VictorÎncă nu există evaluări
- Principiul Minimului de Energie Si Maximul de EntropieDocument9 paginiPrincipiul Minimului de Energie Si Maximul de EntropieCristina MoldoveanuÎncă nu există evaluări
- Introducere Curenti TurbionariDocument3 paginiIntroducere Curenti TurbionariEu VictorÎncă nu există evaluări
- Determinarea Rezistivitatiii de VolumDocument4 paginiDeterminarea Rezistivitatiii de VolumDan Petre100% (1)
- Radar ComponenteDocument2 paginiRadar ComponenteEu VictorÎncă nu există evaluări
- Determinarea Rezistivitatiii de VolumDocument4 paginiDeterminarea Rezistivitatiii de VolumDan Petre100% (1)
- Linii Electrice SubteraneDocument5 paginiLinii Electrice SubteraneEu VictorÎncă nu există evaluări
- Linii Electrice SubteraneDocument5 paginiLinii Electrice SubteraneEu VictorÎncă nu există evaluări
- Fiziopatologia InfectieiDocument6 paginiFiziopatologia InfectieiRoxanaktÎncă nu există evaluări
- Catabolismul AminoacizilorDocument2 paginiCatabolismul AminoacizilorEu VictorÎncă nu există evaluări