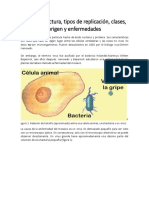
S-ar putea să vă placă și
- Infografía Tipos de ReproducciónDocument6 paginiInfografía Tipos de ReproducciónEdson Noel Flórez Alvarado100% (1)
- Farmacologia BasicaDocument171 paginiFarmacologia BasicaAntonio Jesús MartinzÎncă nu există evaluări
- Generalidades VirusDocument38 paginiGeneralidades VirusMateo Geovo Correa100% (1)
- Por Qué Se Debate Si Los Virus Son o No Seres VivosDocument9 paginiPor Qué Se Debate Si Los Virus Son o No Seres VivosVictor GarciaÎncă nu există evaluări
- Genetica BacterianaDocument9 paginiGenetica BacterianaLiz GarciaÎncă nu există evaluări
- Mecanismos de Replicación de ProcariotasDocument3 paginiMecanismos de Replicación de Procariotasfernando k3Încă nu există evaluări
- Los VirusDocument29 paginiLos Virusclaudia agudo100% (1)
- Los VirusDocument9 paginiLos VirusCyberÎncă nu există evaluări
- Origen de Los VirusDocument3 paginiOrigen de Los VirusItalo A. Figueroa50% (2)
- El Experimento de Hershey FARMADocument8 paginiEl Experimento de Hershey FARMAJesus Federico Rios VelazquezÎncă nu există evaluări
- ¿Están Vivos Los Virus - Un Nuevo Descubrimiento Indica Que Se Parecen Más A Los Zombis - Ciencia - EL PAÍSDocument5 pagini¿Están Vivos Los Virus - Un Nuevo Descubrimiento Indica Que Se Parecen Más A Los Zombis - Ciencia - EL PAÍSmejocoba82Încă nu există evaluări
- El Tipo de ReproducciónDocument3 paginiEl Tipo de ReproducciónSOFI FERNANDEZÎncă nu există evaluări
- Biología y Genética - Mecanismos de Transferencia NaturalDocument26 paginiBiología y Genética - Mecanismos de Transferencia NaturalMirka jordanÎncă nu există evaluări
- Parcial de BiologiaDocument4 paginiParcial de BiologiaJoalimar Ariana Villegas RequenaÎncă nu există evaluări
- Combinación GeneticaDocument11 paginiCombinación GeneticaAriana RojasÎncă nu există evaluări
- TeatroDocument10 paginiTeatroabigailvizcaya9Încă nu există evaluări
- La Reproduccion BacterianaDocument4 paginiLa Reproduccion BacterianayamilkaÎncă nu există evaluări
- 4to TallerDocument6 pagini4to TallerJesusVasquezLuzardoÎncă nu există evaluări
- Mecanismos de ReproducciónDocument8 paginiMecanismos de ReproducciónSofia castroÎncă nu există evaluări
- Evaluacion Reproduccion Parte 1Document1 paginăEvaluacion Reproduccion Parte 1juanb83Încă nu există evaluări
- Reproducción AsexualDocument2 paginiReproducción AsexualIvan RochinÎncă nu există evaluări
- Los VirusDocument4 paginiLos VirusLore Cecilia VelasÎncă nu există evaluări
- Señalización Célula A Célula en Organismos UnicelularesDocument5 paginiSeñalización Célula A Célula en Organismos UnicelularesOscar VillanuevaÎncă nu există evaluări
- Genetica BacterianaDocument9 paginiGenetica BacterianaMizael AmiquechuÎncă nu există evaluări
- (Spanish) The Pandemic That Lasted 15 Million Years (DownSub - Com)Document11 pagini(Spanish) The Pandemic That Lasted 15 Million Years (DownSub - Com)Luis Mariano Soldevilla ChoqueÎncă nu există evaluări
- Lectura VirusDocument4 paginiLectura VirusLuis Fernando Martínez SantoyoÎncă nu există evaluări
- Juliana Niño Salamanca 803 20 de Marzo Del 2020Document7 paginiJuliana Niño Salamanca 803 20 de Marzo Del 2020Alexandra SalamancaÎncă nu există evaluări
- Tienen Vida Los VirusDocument4 paginiTienen Vida Los VirusCynthia Ponce de LeonÎncă nu există evaluări
- Virus y BacteriofagosDocument11 paginiVirus y Bacteriofagositachi_11Încă nu există evaluări
- Mari Pope OsborneDocument5 paginiMari Pope OsborneCaleb Santiago Forero OteroÎncă nu există evaluări
- Material Genético 4to MedioDocument9 paginiMaterial Genético 4to MedioHumberto C. BustosÎncă nu există evaluări
- Recombinación Genética PDFDocument5 paginiRecombinación Genética PDFJavan ZaldivarÎncă nu există evaluări
- El Origen de La Celula EucariotaDocument5 paginiEl Origen de La Celula EucariotaGuillermo SarabiaÎncă nu există evaluări
- Tienen Vida Los Virus IyC Feb05 PDFDocument3 paginiTienen Vida Los Virus IyC Feb05 PDFs3rgio7Încă nu există evaluări
- Vibriones, Priones y VirusDocument5 paginiVibriones, Priones y VirusEMILY VILLARREALÎncă nu există evaluări
- Texto 4 Transducción Bacteriana - En.esDocument7 paginiTexto 4 Transducción Bacteriana - En.esPretty LitiñaÎncă nu există evaluări
- Introducción A La VirologíaDocument19 paginiIntroducción A La VirologíaSergio OrtegaÎncă nu există evaluări
- ResumenDocument41 paginiResumenSebas MansillaÎncă nu există evaluări
- ¿Están Vivos Los Virus? Un Nuevo Descubrimiento Indica Que Se Parecen Más A Los ZombisDocument3 pagini¿Están Vivos Los Virus? Un Nuevo Descubrimiento Indica Que Se Parecen Más A Los ZombisRocio MaidanaÎncă nu există evaluări
- Infografía de Los Tipos de Reproducción en Los Seres VivosDocument14 paginiInfografía de Los Tipos de Reproducción en Los Seres VivosDerek ToddÎncă nu există evaluări
- Guia deREPRODUCCION EN PROTISTOS, BACTERIAS Y HONGOS. Octavo. 2. Primer PeriodoDocument2 paginiGuia deREPRODUCCION EN PROTISTOS, BACTERIAS Y HONGOS. Octavo. 2. Primer PeriodoAngie nataly TiqueÎncă nu există evaluări
- GUÍA 4 BIOLOGIA Aoctavo 2 Reproduccion en Bacterias Protistas y Hongos.Document6 paginiGUÍA 4 BIOLOGIA Aoctavo 2 Reproduccion en Bacterias Protistas y Hongos.jose valderrama100% (1)
- El Origen de La Celula EucariotaDocument5 paginiEl Origen de La Celula EucariotaRosamel DelgadoÎncă nu există evaluări
- Guà - A 1°EVAL. LAB - VIRO 12224Document10 paginiGuà - A 1°EVAL. LAB - VIRO 122247 Esmeralda Galicia CondeÎncă nu există evaluări
- Virus 8°Document9 paginiVirus 8°JHONNY NAVARROÎncă nu există evaluări
- Resumen IBMC (Clase 10 A 11)Document33 paginiResumen IBMC (Clase 10 A 11)Flor GonzalezÎncă nu există evaluări
- La EvolucionDocument5 paginiLa Evolucionvaleria fernandezÎncă nu există evaluări
- VirologìaDocument24 paginiVirologìaMauro Esteban Carvajal CarvajalÎncă nu există evaluări
- Universidad Nacional San Luis Gonzaga de Ica: Facultad de Agronomía Curso: DocenteDocument25 paginiUniversidad Nacional San Luis Gonzaga de Ica: Facultad de Agronomía Curso: DocenteGKCÎncă nu există evaluări
- Vibriones, Priones y VirusDocument5 paginiVibriones, Priones y VirusEMILY VILLARREALÎncă nu există evaluări
- Guia O1 JN 2019Document2 paginiGuia O1 JN 2019Patricia MartínezÎncă nu există evaluări
- Apuntes Biología 3er ParcialDocument21 paginiApuntes Biología 3er ParcialAlessandra BernardoÎncă nu există evaluări
- Informe Sobre Las EubacteriasDocument18 paginiInforme Sobre Las EubacteriasKatherine QuirozÎncă nu există evaluări
- Definir La Vida. Los Virus.Document8 paginiDefinir La Vida. Los Virus.Brenda BernalÎncă nu există evaluări
- Sexual y Asexual BacterianaDocument9 paginiSexual y Asexual BacterianaJuan Carlos FigueroaÎncă nu există evaluări
- Virus BiologiaDocument8 paginiVirus BiologiaPedroÎncă nu există evaluări
- Los Virus Campeones de La EvolucionDocument6 paginiLos Virus Campeones de La EvolucionPeGaZuS911 GaMePLaY y Animacion Florez100% (1)
- Resumen Genética MicrobianaDocument7 paginiResumen Genética MicrobianaHuitzilihut Cardenas OrtegaÎncă nu există evaluări
- T2. Biología MolecularDocument15 paginiT2. Biología MolecularPaula GallegoÎncă nu există evaluări
- Características Esenciales de VirusDocument3 paginiCaracterísticas Esenciales de VirusMathias Lira CornejoÎncă nu există evaluări
- El Fenómeno BullyingDocument15 paginiEl Fenómeno BullyingPamelaÎncă nu există evaluări
- Resumen OrganulosCelularesDocument2 paginiResumen OrganulosCelularesPamelaÎncă nu există evaluări
- Tema 1Document7 paginiTema 1PamelaÎncă nu există evaluări
- AfasiaDocument21 paginiAfasiaPamelaÎncă nu există evaluări
- Adicción A Drogas y GestaciónDocument14 paginiAdicción A Drogas y GestaciónPamelaÎncă nu există evaluări
- Fisiologia VegetalDocument91 paginiFisiologia VegetalPamelaÎncă nu există evaluări
- Glaciares en ChileDocument66 paginiGlaciares en ChilePamela100% (4)
- Articulo Cientifico Sobre El Sistema Nervioso PDFDocument6 paginiArticulo Cientifico Sobre El Sistema Nervioso PDFJenni Mendocilla100% (1)
- Reparacion TisularDocument35 paginiReparacion TisularJaime Andrés Bravo Tapia100% (1)
- Activacion PLaquetariaDocument18 paginiActivacion PLaquetariaJoás Daniel MamaniÎncă nu există evaluări
- Vías Inflamatorias en La Resistencia A La InsulinaDocument4 paginiVías Inflamatorias en La Resistencia A La InsulinaRuth Vargas ChaÎncă nu există evaluări
- Nuestros Desconocidos Cuatro Cerebros - Una Verdad Jamás ImaginadaDocument7 paginiNuestros Desconocidos Cuatro Cerebros - Una Verdad Jamás Imaginadabladimir mazaÎncă nu există evaluări
- Señalizacion CelularDocument25 paginiSeñalizacion CelularJessenia TaipeÎncă nu există evaluări
- Práctica 1 Receptores SensorialesDocument14 paginiPráctica 1 Receptores SensorialesDaniel James Arriaga80% (5)
- Mecanismos de Acción Hormonal - 4 AnimacionesDocument5 paginiMecanismos de Acción Hormonal - 4 AnimacionesHarold KnudsonÎncă nu există evaluări
- Efectos de La Radiacion UvB en Chenopodium Quinoa PDF 2018.en - EsDocument12 paginiEfectos de La Radiacion UvB en Chenopodium Quinoa PDF 2018.en - EspolÎncă nu există evaluări
- Guía 1 GlicobiologíaDocument12 paginiGuía 1 GlicobiologíaSandra R. Manrique HernándezÎncă nu există evaluări
- SC 1Document28 paginiSC 1Justina GirautÎncă nu există evaluări
- Caso Parecido A SinjinDocument38 paginiCaso Parecido A SinjinRODRIGO SABA COSSIOÎncă nu există evaluări
- Comunicación IntercelularDocument10 paginiComunicación IntercelularthiagotavoraÎncă nu există evaluări
- Introducción A La Regulación y La Señalización MolecularesDocument6 paginiIntroducción A La Regulación y La Señalización MolecularesmontseÎncă nu există evaluări
- Patologuia SeminarioDocument28 paginiPatologuia SeminarioJennifer Navarro GarciaÎncă nu există evaluări
- Receptores Con Actividad EnzimáticaDocument25 paginiReceptores Con Actividad EnzimáticaNicolas Ignacio Aliaga MachucaÎncă nu există evaluări
- Biofarmacos en EspanaDocument11 paginiBiofarmacos en EspanaAnaÎncă nu există evaluări
- TransduccionDocument53 paginiTransduccionJonatan David Escalante ORtizÎncă nu există evaluări
- Discusión de Grupo Nº4 PDFDocument20 paginiDiscusión de Grupo Nº4 PDFJuly CamposÎncă nu există evaluări
- La Transmicion NerviosaDocument7 paginiLa Transmicion Nerviosaeduardo100% (2)
- Tarea 7 - Realizar Análisis de Artículo - Unidad 1 - Unidad 2 - Unidad 3 - Erika - ForeroDocument8 paginiTarea 7 - Realizar Análisis de Artículo - Unidad 1 - Unidad 2 - Unidad 3 - Erika - Foreroerika forero100% (1)
- 15.1 Los Elementos Básicos de Los Sistemas de Señalización CelularDocument6 pagini15.1 Los Elementos Básicos de Los Sistemas de Señalización CelularALEJANDRA INÉS NIETO ARIASÎncă nu există evaluări
- Efectos Dirigidos y No Dirigidos de La Radiación IonizanteDocument12 paginiEfectos Dirigidos y No Dirigidos de La Radiación IonizantePaola12Încă nu există evaluări
- Guia GemfinDocument42 paginiGuia Gemfin82lauÎncă nu există evaluări
- Tumores Hipotalámicos, Hipofisiarios y SillaresDocument15 paginiTumores Hipotalámicos, Hipofisiarios y SillaresFridaVianelPat100% (1)
- Tesis Miryam Ochoa Agredano - 24 Feb 2022Document61 paginiTesis Miryam Ochoa Agredano - 24 Feb 2022Augusto RodriguezÎncă nu există evaluări
- Farmacologi A1Document131 paginiFarmacologi A1Pedro MartínezÎncă nu există evaluări
- Dosis Respuesta Seminario 1Document24 paginiDosis Respuesta Seminario 1Andy AlvaradoÎncă nu există evaluări
- Alto Rendimiento Deportivo M3T9Document7 paginiAlto Rendimiento Deportivo M3T9lautaroÎncă nu există evaluări