Documente Academic
Documente Profesional
Documente Cultură
Semnalizarea Celulara
Încărcat de
Ovidiu VaidaTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Semnalizarea Celulara
Încărcat de
Ovidiu VaidaDrepturi de autor:
Formate disponibile
SEMNALIZAREA
CELULARA
Toate organismele au la baz urmtoarele aciuni fundamentale:
obinerea nutrienilor din mediu i eliminarea produselor de degradare
obinerea energiei prin oxidarea compuilor chimici
reproducere i cretere
micare
rspuns la mediul nconjurtor
La organismele unicelulare (procariote) fiecare celul i realizeaz
singur aceste aciuni.
n organismele multicelulare, fiecare funcie este ndeplinit de
celule specializate.
Aceasta specializare necesit n schimb coordonarea activitilor
esuturilor specializate, proces realizat prin mecanisme de
semnalizare intercelular.
Controlul alosteric al metabolismului, care apare n toate celulele,
procariote i eucariote, este tipul de modulare a activitii enzimatice
n care metaboliii cilor biochimice controleaz prin feed-back
enzimele, pe aceleai ci sau pe ci diferite
control n ntregime intracelular
nu pot lua decizia ce cale metabolic ar trebui urmat
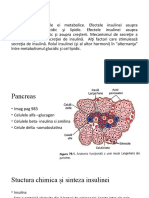
de ex. o celul hepatic poate s stocheze glicogenul sau s-l
consume. Celula hepatic, ea nsi, nu poate decide aceasta,
procesul fiind determinat de semnalele chimice produse de alte
celule, n exemplul folosit, de insulin i glucagon, produse de
pancreas.
Controlul metabolic este numai vrful iceberg-ului semnalelor
celulare.
Procesele celulare fundamentale sunt determinate de semnalul
extracelular.
Dac semnalele primite nu sunt acceptate, celulele normale
(necanceroase) nu cresc, nu se divid i nu se difereniaz.
n cazul celulelor canceroase, cile semnal opereaz ntr-un mod
anormal, astfel nct celulele se divid necontrolat.
Faptul c semnalele extracelulare controleaz multe activiti vitale
n numeroase celule diferite, susine potenialitatea interveniei
terapeutice n multe dintre boli, numeroase medicamente fiind
orientate spre semnalele celulare.
Elementele de baz ale semnalizrii
celul semnalizatoare elibereaz semnalul
ligand - molecul semnal eliberat de celula semnalizatoare
celule int - celule echipate pentru captarea semnalului
Molecul semnal extracelular
Receptor proteic
Cale de semnalizare intracelular
Protein reglatoare Proteine int a unei protein a
a unei gene enzime metabolice citoscheletului
modificarea expresiei metabolism modificarea formei
genice modificat sau micrii celulare
celula int dispune de receptori proteici specifici la care se leag
moleculele semnal, un rspuns biochimic n interiorul celulei.
receptorii proteici specifici - pe suprafaa sau n interiorul celulei int.
transducia (interpretarea) semnalului = mecanismele prin care
semnalele sunt recepionate, interpretate i cile prin care
mainria celular rspunde la aceste semnale. (contracie,
cretere, secreie, modificri metabolice)
Rspunsurile celulare includ:
modularea expresiei genelor specifice
efecte metabolice directe.
Fiecare tip de celul este capabil s comunice = capacitatea de a
produce, recunoate, interpreta i rspunde la semnalele din
mediul nconjurtor (snge, limf, celule vecine).
Majoritatea semnalelor sunt molecule chimice
Componentele celulare de transfer al semnalului = elemente de
traducere ale semnalului n limbajul celulei.
Rspunsul celular la semnal
ntre semnalul extern i efectele celulare au loc evenimente n cascad
= cascad de semnalizare.
Simplificat, toate cascadele de semnalizare au obligatoriu urmtoarele
etape:
ligant (semnal) receptor mesager efect celular
Rspunsurile celulare necesit:
milisecunde (exemplu contracia muscular indus pe cale
nervoas) implic reglarea numai a proteinelor existente
ore, zile, luni (rspunsurile la semnale hormonale) implic
modificri n tipul i cantitatea proteinelor.
Strategii de semnalizare
Semnalizare la distan. Un exemplu n acest sens este aciunea
hormonal de tip endocrin, paracrin sau autocrin.
Semnalizare de contact. Realizat n cadrul adeziunii celulare, ntre
2 celule vecine sau ntre celul i matricea celular. Rspunsurile
celulare au loc n urma modificrilor contactului celulei cu alte celule
(adeziunea celular) sau cu matricea celular.
Semnalizare direct. De exemplu, molecule semnal din citoplasma
unei celule ajung n citoplasma celulei vecine prin canale de
jonciune.
Principalele ci de transfer intracelular
ale informaiei sunt:
Modificarea funcionrii unor canale ionice transmembranare pentru
Ca+2, K+, Na+, Cl-
Modificarea activitii unor receptori enzime transmembranare de
exemplu receptorii cu activitate tirozin-kinazic sau sisteme JAK-STAT.
S Ioni
S
Canal deschis
Canal nchis
Membran
S
Substrat-Tyr Substrat-Tyr- P
S
Membran
Modificarea conformaiei unor receptori transmembranari cu 7
domenii alfa -helix ce induc activarea unor enzime productoare de
semnale intracelulare (mesageri secunzi). De exemplu:
Activarea transcripiei unor gene specifice prin legarea de
receptori intracelulari, de exemplu semnalizarea prin hormoni
hidrofobi.
G
Enzim
Mesager
secund
S
G
S
Membran
R
R
S ARNm
proteine
Membran
S
Modificarea conformaiei citoscheletului celular prin
intermediul receptorilor de adenozin ce leag molecule din
matricea extracelular.
n jur de 30% din cele aproximativ 30.000 de gene umane codific
sinteza de proteine implicate n cile semnal: receptori, proteine
G, enzime productoare de mesageri secunzi, protein kinaze,
proteine de desensibilizare, proteine de tip canal ionic. Aceste
proteine prezint o organizare domenial cu elemente specifice
comune (SH2, SH3, domenii de legare ADN, domeniu Tyr-P etc)
domenii conservate de-a lungul evoluiei la majoritatea speciilor.
membran
Fibr
citoschelet
Tipuri de semnale
Semnalele celulare = molecule chimice organice sau anorganice ce apar
n mediul nconjurtor.
Numrul acestora este de ordinal sutelor, i se descoper altele, practic n
fiecare zi.
Pot fi proteine, peptide mari, peptide mici, aminoacizi, nucleotide,
steroizi, eicosanoizi, hormoni tiroidieni, epinefrine, derivai ai
vitaminelor A i D, gaze (oxid nitric)
Structura lor chimic nu are relevan pentru mecanismul aciunii lor, ea
interesnd numai n legarea cu receptorii celulari specifici.
Legtura dintre molecula semnal i receptorul su specific este o
legtur necovalent, foarte specific i reversibil.
Molecul semnal = orice molecul ce induce un rspuns specific la
nivelul unei celule int.
1. endocrini = hormoni, produi de glande endocrine, ce acioneaz la
distan de celulele secretoare
2. mediatori locali sau paracrini - acioneaz local asupra celulelor vecine
(ex. transmiterea impulsului nervos); factorii de reglare ai inflamaiei la
nivelul unui sector infecie.
3. autocrini - anumite celule rspund la molecule semnal pe care le produc
ele nsele (ex. limfocitele T rspund la stimularea antigenic prin sinteza
unui factor de cretere care determin autoproliferarea lor)
4. contactul fizic intre celule vecine care va genera un semnal contact-
dependent.
Moleculele semnal hidrosolubile - neurotransmitori, hormoni derivai de
AA, hormonii proteici, factorii de cretere proteici, nu pot strbate direct
stratul dublu lipidic al membranei celulare. Ele se leag de domeniul
exterior al receptorului membranar mesager secund.
Moleculele semnal liposolubile - hormonii steroizi, hormonul tiroidian,
acidul retinoic, retinoizii, vitamina D3, intr direct n celul i se leag de
receptorii intracelulari (nucleari i citoplasmatici) ai celulei int.
Protein
kinaze
Receptor
Hormon
Mesageri secunzi
mARN
P
Gene
NUCLEU
CITOPLASM
Aciunea receptorilor de membran Aciunea receptorilor nucleari
Hormon
Receptor
mARN
CITOPLASM
NUCLEU
Gene
P
Rspunsul la molecule semnal mediat de receptorii intracelulari
1. receptori citoplasmatici - legarea H la R modificri
conformaionale n complexul steroid-receptor (SR), fenomen numit
activare sau transformare.
Activarea/transformarea conduce la translocarea SR n nucleu unde
acest complex interacioneaz cu ADN i cromatina pentru a
influena sinteza proteic.
2. Receptorii nucleari au un domeniu central de legare la ADN
(DBD) care orienteaz receptorul spre secvenele specifice ADN,
cunoscute ca elemente de rspuns hormonal.
DBD este compus din 2 regiuni nalt conservate zinc finger care
difereniaz receptorii nucleari de alte proteine de legare la ADN.
LBD poate fi imaginat simplu ca un comutator molecular care dup
legarea ligandului transform receptorul ntr-o stare transcripional
activ.
Superfamilia receptorilor nucleari :
familia receptorilor steroidici i
familia receptorilor tiroidieni/retinoizi/vitamina D.
Fiecare tip de receptor are subfamilii (de exemplu subfamilia
receptorilor pentru acid retinoic). Aceste subtipuri sunt produse ale
unor gene individuale (RAR, RAR, RAR) iar izoformele
receptorilor apar datorit splicing-ului difereniat i/sau utilizarea
unor promotori diferii (RAR1, RAR2).
Un receptor nuclear tipic conine o regiune variabil N-terminal
(A/B), una conservat DBD (C), o regiune median variabil (D),
una conservat LBD (E) i o regiune variabil C-terminal.
Receptorii nucleari pot fi mprii n 4 clase funcie de legarea
liganzilor, a ADN i proprietile de dimerizare:
receptori steroidici,
heterodimeri RXR,
receptori homodimeri orfani,
receptori monomeri orfani.
Pentru receptorii orfani nc nu se cunosc liganzii naturali.
N C
DNA Ligand
A/B C D E F
GR GR
GR
MR
PR
AR
ER
glucocorticoid
mineralcorticoid
progesteron
androgen
estrogen
TR
RAR
VDR
PPAR
EcR
hormon tiroidian
all-trans RA
1,25-(OH)
2
-VD
3
eicosinoide
ecdison
RXR RAR
Receptori steroidici Heterodimeri RXR
Receptori dimeri orfani
RXR (usp)
COUP/ARP
HNF-4
9-cis RA
?
?
Receptori monomeri orfani
?
?
RXR RXR
NGFI-B
NGFI-B
ELP/SF-1
DBD LBD
N C
DNA Ligand
A/B C D E F
GR GR
GR
MR
PR
AR
ER
glucocorticoid
mineralcorticoid
progesteron
androgen
estrogen
TR
RAR
VDR
PPAR
EcR
hormon tiroidian
all-trans RA
1,25-(OH)
2
-VD
3
eicosinoide
ecdison
RXR RAR
Receptori steroidici Heterodimeri RXR
Receptori dimeri orfani
RXR (usp)
COUP/ARP
HNF-4
9-cis RA
?
?
Receptori monomeri orfani
?
?
RXR RXR
NGFI-B
NGFI-B
ELP/SF-1
ARE
Dimerizare
Translocare
Disociere HSP
FKBP52
HSP90
HSP70
DHT
5reductaz
Testosteron
Citosol
Nucleu
P
P P P
Fosforilare
Rspuns celular
CoA
CBP
ARNpolI
Calea semnal atestosteronului
Membrana nuclear
Element de legare la ADN
ADN
CITOPLASM
NUCLEU
receptor
HSP
Glucocorticoid
Por
nuclear
Domeniu de legare la ADN
Activarea transcripiei
Calea semnal a glucocorticoizilor
Rspunsul la molecule semnal mediat de receptorii
membranari
Majoritatea cilor semnal utilizeaz receptorii membranari.
Receptori membranari
proteine specifice localizate pe suprafaa celulelor care acioneaz
ca transductori ai semnalului transportat de o molecul ligant;
leag ligandul cu mare afinitate i transform acest semnal
extracelular ntr-un semnal intracelular care modific
comportamentul celulei int.
are un domeniu extern la care se leag molecula semnal, un
domeniu transmembranar i un domeniu citoplasmatic.
Cnd molecula semnal se leag de receptorul su specific,
domeniul su citoplasmatic sufer o modificare conformaional
(prin adugarea unui grup fosforil) care iniiaz calea semnal
Calea semnal - finalitate - activarea de gene specifice i/sau
sisteme metabolice de control.
Cheia procesului de transmitere a semnalului celular este
fosforilarea intermediarilor proteici de semnalizare de ctre
proteinkinaze i defosforilarea de ctre proteinfosfataze.
Calea RAS
TK
TK
MEK
ERK
Calea PI-3
kinaza
TK
Fosfatidil-
inozitol kinaza
Fosfatidil-
inozitol(3,4,5)P
3
PKB
Glicogen
sintaza
Efecte
celulare
Calea
JAK/
STAT
JAK
Proteine
STAT
G G G
Calea
cAMP
Calea
cGMP
Lumin
Calea
Fosfatidil
- inozitol
cAMP
PKA
PLC
Efecte
celulare
DAG
IP
3
Ca
2+
PKC
Efecte
celulare
(cGMP)
Vedere
cGMP
Efecte
celulare
NO
cGMP
Efecte
celulare
Strategii de semnalizare
Clasificarea tipurilor de receptori membranari
1. receptorii tirozin-kinazici - enzime, activitatea le permite
asocierea cu molecule adaptor ce iniiaz calea semnal.
Funcie de tipul moleculei adaptor se iniiaz urmtoarele cai semnal:
calea Ras
calea fosfatidil-inozit 3-kinaza (PI 3)
calea receptorilor JAK/STAT
Legtura moleculei semnal determin dimerizarea receptorilor, urmat de
autofosforilarea gruprilor tirozin ale domeniilor lor citoplasmatice, acestea
avnd ele nsele activitate tirozin-kinazic.
n citoplasm exist proteine care se leag la receptorii fosforilai i
acioneaz ca molecule adaptor - legtura dintre receptorii activai i cile
semnal care transmit semnalul de cele mai multe ori la nucleu.
n apropierea locusului fosforilat al unui receptor exist secvene de
aminoacizi la care aceste proteine se leag specific, astfel c adaptorul
proteic corespunztor se leag la receptorul corespunztor.
La rndul lor, proteinele adaptor au locusul propriu de legare specific.
2. receptorii asociai cu proteine G - acioneaz indirect,
reglnd activitatea unei alte proteine legat de membrana citoplasmatic,
proteina ce poate fi o enzim sau un canal ionic.
Interaciunea dintre receptori i proteina int este mediat de o a treia
protein trimeric, legat de GDP sau GTP (proteina G).
Acesti receptori nu dimerizeaz i nu sunt fosforilai.
Legarea moleculei semnal cauzeaz o modificare
conformaional a domeniului citoplasmatic al receptorului.
Au de asemenea molecule adaptor care sufer prin legare
modificri conformaionale i leag receptorul la calea semnal
Cile semnal iniiate prin acest mecanism sunt :
calea AMPc
calea GMPc
calea fosfatidilinozitol
procesul de transmitere a vederii calea transmiterii energiei
luminoase
procesul de transmitere a gustului, mirosului celule
senzoriale specifice
.
Efecte celulare
(a)
P
P
P
P
P
P
P
P
Membrana citoplasmatic
Tirozinkinaz
Protein citoplsmatic
ataat la receptorul
fosforilat (adaptor)
Domeniul de legtur
SH
2
Efecte celulare
(b)
Calea Ras
Receptor tirozin kinazic
implic receptorul i o cascad de 3 proteinkinaze citoplasmatice
(Raf, MEK, ERK MAP kinaze), fiecare activnd prin fosforilare
urmtorul component al cii.
La finalul cascadei, ultimul element (ERK activat) ptrunde n nucleu
unde, prin fosforilarea unor factori transcripionali, activeaz gene
specifice.
n aceast cale nu exist molecule mici cu rol de mesageri secunzi,
toate componentele sunt proteine.
Exist peste 50 de receptori cunoscui:
factorul de cretere epidermal (EGF),
factorul de cretere derivat din plachete (PDGF),
factorul de cretere fibroblastic (FGF),
factorul de cretere hepatocitar,
insulina,
factorul de cretere al endoteliului vascular (VEGF).
Membrana
celular
Receptor inactiv EGF
citoplasm
GRB SOS
Ras
Raf
GDP
P P P P GTP
Raf GRB SOS
Ras
MEK
P P
ERK
P P
Por nuclear
P P
ERK
P P
Factori de
transcripie inactivi
Factori de
transcripie activai
P P
Secven reglatoare a
genei
Transcripie
genic
nucleu
Mecanismul comutator Ras GTP/GDP
Ras deine un mecanism comutator de tip
nchis/deschis utilizat n mare msur n controlul cilor
semnal. Ras aparine unei familii de proteine numite
monomeri mici GTP-azici. Au legat o molecul de GDP
sau GTP. Ras este activ n forma GTP i inactiv n
forma GDP (figura 14).
Pentru activarea Ras (proces n urma cruia
aceasta activeaz urmtorul component) GDP trebuie
schimbat cu GTP, aceast schimbare avnd loc numai
cnd Ras este n contact cu complexul GRB-SOS legat
la receptorul activat (n cazul de fa, numai cnd EGF
este ataat la receptor). n urma schimbrii GDP cu
GTP, proteina Ras-GTP trece pe poziia deschis i
induce activarea urmtorului element al cii. Aceast
stare activat dureaz foarte puin deoarece Ras,
posednd capacitate GTP-azic, hidrolizeaz rapid GTP,
revenind n starea inactiv Ras-GDP.
Inactivarea i modularea cii Ras
Ras este inactivat de activitatea sa intrinsec GTP-azic. MEK i ERK
activate sunt inactivate de protein-fosfataze specifice prin defosforilarea
lor. A fost descoperit c proteinkinazele i fosfatazele corespunztoare sunt
fizic legate mpreun, sugernd faptul c timpii n care proteina este sub
forma fosforilat respectiv defosforilat se succed foarte rapid, proteina
trecnd prin strile activ inactiv.
O cale semnal scpat de sub control este caracteristic pentru
celulele canceroase.
Exist multiple ci similare de tipul Ras, fiecare cu o cascad de
proteinkinaze terminat cu o MAP-kinaz care ptrunde n nucleu i
activeaz factorii transcripionali int.
Sunt astfel multiple ci de tip Ras opernd n paralel, cu proteinkinaze
specifice corespunznd ca funcie la Raf, MEK i ERK i cu diferii factori
transcripionali int. Acestea nu apar ca i ci liniare izolate, ntre ele
existnd interconexiuni.
Aceste ci multiple pot fi considerate curieri ai semnalelor de la diferii
receptori la nucleu unde activeaz gene specifice fiecrui tip de semnal.
Nu se cunoate cum receptori complet diferii pot activa aceiai cale
Ras dar rspunsurile celulare la cele dou semnale sunt complet diferite.
Este cunoscut c gruprile kinazelor din diferitele ci sunt legate mpreun
prin proteinele schel (scaffold proteins) care astfel probabil aleg calea.
Cile semnal diferite par s fie interconectate ntre ele, element esenial n
coordonarea multiplelor semnale pe care celula le primete.
Calea PI 3-kinaza (fosfatidil-inozit-3-kinaza)
controlul proliferrii i diferenierii celulare
controlul metabolic
activat de receptori rspunznd la un numr foarte
mare de hormoni (insulina), factori de cretere i
neurotransmitori
form oncogenic a PI3-kinaza.
fosfatidilinozitol 3-kinaza (PI3-kinaza) - Substratul este
un component membranar numit fosfatidilinozitol-4,5-
bifosfatul (PI(4,5)P2) care este fosforilat, rezultnd
PI(3,4,5)P3 sau PIP3 (fosfatidil inozitol trifosfat) -
mesager secund.
PIP3 activeaz proteinkinaze sau alte enzime
Receptor insulinic
IRS
PI3-kinaza
PI(4,5)P
2
insulina
P
P
P P
IRS
fosforilat
PI3-kinza
activat
PI(3,4,5)P
3
Mesag. sec.
kinaz
PKB-P
activat
Fosfataza Glicogen-
sintetazei activat
La legarea insulinei, domeniul citoplasmatic al receptorului, care
este o tirozinkinaz, devine fosforilat la gruparea hidroxil ale
resturilor de tirozina.
Proteina adaptor IRS se leag la acest receptor fosforilat i este la
rndul su fosforilat de tirozinkinaza receptorului.
O enzim numit fosfatidilinozitol 3-kinaza (PI3-kinaza) se leag la
IRS fosforilat, activndu-se n acest fel i apropiindu-se de
membrana citoplasmatic unde se gsete substratul PI3-kinazei,
un component membranar numit fosfatidilinozitol-4,5-bifosfatul
(PI(4,5)P2). Sub aciunea PI3-kinazei, acesta este fosforilat,
rezultnd PI(3,4,5)P3 sau PIP3 (fosfatidil inozitol trifosfat), ce
constituie mesagerul secund.
PIP3 atrage PKB citoplasmatic inactiv, atand-o la membran.
O kinaz localizat membranar (PDK1) determin activarea
proteinkinazei B (PKB) prin fosforilare
PKB activat determin activarea prin fosforilare a fosfatazei
glicogen sintetazei fosforilate inactive i prin urmare se stimuleaz
sinteza de glicogen.
PKB activeaz i transferul intracelular al glucozei (GLUT4)
P
IRS-1
P
P
PI-3K
P
GS
(inactiv)
GS
(activ)
GSK3
(inactiv)
GSK3
(activ)
P
PKB
P
P
GLICOGEN
GLUT4
Glucoz
PIP
2
PIP
3
Conexiuni ntre calea Ras i calea PI3-kinazei
Se constat c acelai semnal, de exemplu insulina poate declana
concomitent 2 ci de semnalizare diferite, calea PI3-kinazei i calea Ras
MAPK.
Este o cale mediat prin tirozin kinaze cu deosebirea c nu receptorul posed
activitate tirozin kinazic ci o tirozin kinaz solubil din citoplasm ce se
ataeaz de receptorul modificat n urma legrii semnalului.
Calea este utilizat de citokine, eritropoietin, interferon, etc.
Raf-1
Receptor insulinic
ERK
Ras
Sos
Grb2
P
P
MEK
MP1
P
P
IRS-1
P
P
PKC
PKB
PI-3K
PIP
3
PIP
3
14-3-3
Calea JAK/STAT
Interferonii, proteine protectoare, eliberate de celulele infectate de virusuri,
citokine
Receptorul specific are asociat la domeniul su citoplasmatic cte o JAK-
kinaz. Legarea interferonului la receptor determin dimerizarea
receptorilor i activarea JAK-kinazelor.
JAK-kinazele au dou situsuri catalitice
JAK-kinazele activate fosforileaz resturile tirozin ale receptorului dimer.
dou STAT proteine din citosol se leag la receptorul fosforilat
JAK-kinazele, rmnnd legate la receptor, fosforileaz cele 2 STAT-
proteine.
Proteinele STAT fosforilate migreaz n nucleu, unde mpreun cu o a 3-a
component proteic GAS (-interferon activates sequence element) se
asambleaz ntr-un factor transcripional activ determin transcripia genelor
specifice.
Spre deosebire de receptorul EGF, care este o tirozin-kinaz, n aceast
cale dimerizarea receptorului activeaz tirozin-kinazele citoplasmatice
(Janus kinazele sau JAK- kinazele).
O trstur caracteristic a acestui tip de cale este numrul redus de
intermediari: factorii citoplasmatici inactivi sunt activai i migreaz n nucleu
unde acioneaz asupra genelor (calea Ras este mult mai complicat).
Receptorii cuplai cu proteina G
superfamilie de receptori folosii de numeroi hormoni i factori de cretere,
neurotransmitori, neuropeptide, rodopsina, activat de lumin, receptorii
olfactivi din mucoasa nazal i receptorii gustativi de la nivelul mugurilor
gustativi.
Jumtate din medicamentele curente au drept inte receptori cuplai cu
proteine G.
Din punct de vedere structural i funcional receptorii sunt proteine nrudite,
cu 7 motive transmembranare, cu structura -helix, motiv pentru care au
fost numii i motivul celor 7 elice. Aceti receptori au un domeniu
extracelular care leag ligandul (ex. proteinele) i un domeniu citoplasmatic
ce face legtura cu proteinele G.
Fiecare receptor are un locus specific citoplasmatic de legare a proteinelor
G heterotrimerice (, i ).
Exist o mare diversitate de proteine G care se pot cupla la o mare
diversitate de ci semnal.
Receptorii asociai proteinei G nu au activitate enzimatic proprie.
Modificarea conformaional a domeniului lor citoplasmatic, care survine n
urma legrii moleculei semnal la domeniul extern al receptorului, cauzeaz
interaciunea domeniului su citosolic cu proteine G asociate.
exemplu tipic: adrenalina activeaz adenilat ciclaza la nivel hepatic, via
proteine G, cu producerea de AMPc i n final glicogenoliza.
receptor -adrenergic: proteina G este ataat la domeniul
citoplasmatic al receptorului. Dac receptorul nu este legat de hormon, la
subunitatea a proteinei G este legat GDP, aceasta este inactiv.
Cnd adrenalina se leag la receptor, aceasta sufer modificri
conformaionale, care determin modificri de structur i la nivelul
proteinei G.
Proteina G, numit n acest caz Gs (s-stimulare), schimb GDP cu
GTP la nivelul subunitii .
Complexul subunitate -GTP se detaeaz de celelalte dou proteine
i , se mic pe faa intern a membranei i se leag de o molecul de
enzim, adenilat-ciclaza pe care o activeaz i care va produce AMPc.
Hormonul determin producia de AMPc folosind proteina G ca intermediar.
Subunitatea a proteinei G, n forma sa GTP, are activitate GTP-
azic, hidroliznd GTP la GDP.
Subunitatea -GDP revine la forma sa iniial, se detaeaz de
adenilat-ciclaz i se reasociaz la complexul , reconstituind proteina G
inactiv. Dac la receptor este legat n continuare un hormon, ntreg ciclul
poate ncepe din nou. Dac receptorul nu mai are un hormon ataat,
procesul nceteaz.
Pentru a continua producia de AMPc, subunitatea a proteinei G
oscileaz ntre receptor i adenilat-ciclaz, durata legrii la adenilat-ciclaz
depinznd de rapiditatea hidrolizei moleculei GTP ataat de subunitatea .
Protein
G
trimeric
Membrana citoplasmatic
Adenilat ciclaza inactiv
Adrenalina
Adenilat ciclaza inactiv
R
GTP
GTP
Adenilat ciclaza activ
AMPc
ATP
GTP
+H
2
O
R
GDP
R
ATP
AMPc
GDP + Pi
R
Receptor
Adenilat ciclaza activ
Adenilat ciclaza inactiv
ATP
GDP
Semnalul epinefrinic (adrenergic)
1. Receptor -adrenergic - Proteina G - Gs (s-stimulare)
2. receptor 2 adrenergic - protein G a crui subunitate -GTP
inhib adenilat-ciclaza, protein G notat Gi (i = inhibiie).
Astfel c un hormon (adrenalina), poate exercita diferite efecte n
diferite esuturi, n funcie de tipul de receptor prezent n celulele
esutului.
n holer - blocarea hidrolizei subunitii -GTP are urmri
dramatice.
Celulele mucoasei intestinale secret Na
+
n lumenul intestinal,
aceast activitate fiind controlat prin intermediul AMPc.
Toxina holeric determin inhibarea activitii GTP-azice a
subunitii a proteinei G. Astfel, odat ce un stimul hormonal
activeaz adenilat-ciclaza, calea se blocheaz n stadiul -GTP.
Prelungirea produciei de AMPc determin pierderi masive de Na
+
i
de ap, determinnd diaree masiv i posibil moarte prin pierderi
hidro-electrolitice.
Efectele induse de AMPc sunt mediate prin activarea, ca urmtor intermediar
a protein kinazei dependente de AMPc (PKA).
n stadiul inactiv, PKA este format din 2 subuniti reglatoare (R) - capabile
s lege fiecare cte 2 molecule de AMPc i 2 subuniti catalitice (C). n
stadiul activ, subunitile reglatoare care au legat 4 molecule de AMPc i
schimb conformaia i se disociaz de unitile catalitice, care fosforileaz
specific moleculele de substrat.
Rspunsuri:
rapide - n celulele musculare striate, PKA activat fosforileaz enzime
implicate n metabolismul glicogenului care declaneaz simultan hidroliza
glicogenului la glucoz i inhib sinteza glicogenului, crescnd cantitatea de
glucoz disponibil n cteva secunde.
lente - PKA activat (de creterea concentraiei AMPc n citosol) elibereaz
subunitile sale catalitice, care se deplaseaz n nucleu, unde fosforileaz
factorul de transcripie numit proteina de legare la elementul de rspuns la
AMPc (CREB). Odat fosforilat, CREB recruteaz o protein coactivatoare
CRB (protein de legare la CREB) i se leag la regiunea reglatoare a
genei (CRE, element de rspuns la AMPc), stimulnd transcripia genei.
Protein-fosfatazele compenseaz efectele proteinkinazelor. Acestea
inactiveaz CREB prin nlturarea fosfatului activ, blocnd rspunsul
transcripional determinat de o cretere a concentraiei AMPc.
Membrana citoplasmatic
adrenalina
Receptor
2
adrenergic
Adenilat ciclaza
activ
GDP
GTP
GTP
ATP
AMPc
Proteina G
R
R
C
C
PKA inactiv
R
R
PKA activat
Membrana nuclear
citosol
Por nuclear
PKA activat
CREB inactiv
P
CRB
CREB fosforilat, activat
Gen int
Element de legare la CREB
TRANCRIPIE
TRANSLAIE
PROTEINE
Oprirea semnalizrii i regenerarea receptorului
Oprirea semnalizrii const n desensibilizarea receptorului
dup activarea produs de moleculele semnal.
Dup legarea adrenalinei de receptor, modificarea conformaiei
acesteia induce i legarea de o proteinkinaz -adrenergic
citoplasmatic (ARK), care fosforileaz receptorul.
Receptorul fosforilat dobndete capacitatea de a lega arestina
(proteina arr). Legarea acesteia detaeaz receptorul de
proteinele G i de membrana plasmatic printr-un proces de
endocitoz, acesta trecnd ntr-o vezicul mic.
Aici este defosforilat i returnat n membrana plasmatic,
redevenind apt (sensibil) la aciunea unei noi molecule de receptor.
Calea cascadei fosfatidilinozitol
(cu proteina G intermediar)
utilizeaz mesageri secunzi provenii din fosfolipidul membranar,
fosfatidilinositol-4,5-difosfat (PIP2).
Aceast cale este utilizat de numeroi hormoni i factori de
cretere.
Legarea hormonului la receptorul specific determin subunitatea
s schimbe GDP cu GTP, apoi se detaeaz i migreaz urmnd s
activeze o enzim legat membranar, fosfolipaza C (PLC) care
hidrolizeaz PIP2 n 2 mesageri secunzi diferii:
inositol-1,4,5-trifosfatul (IP3) i
diacilglicerolul (DAG).
Proteina G hidrolizeaz GTP la GDP i Pi inactivndu-se singur
dup un interval de timp, asemntor cu sistemul adrenergic descris
anterior.
IP3 prsete membrana citoplasmatic i difuzeaz rapid n citosol.
La nivelul reticulului endoplasmatic IP3 se leag la canalele
membranare dependente de IP3, care elibereaz Ca
2+
in citosol.
Ca
2+
eliberat prin canalele deschise va crete rapid concentraia
Ca
2+
din citosol.
Ca
2+
nu se poate lega direct la enzime, ci la o protein receptoare
din citoplasm numit calmodulin (4Ca
2+
) modificri
conformaionale complex activ Ca
2+
/calmodulin.
Ca
2+
/calmodulina activeaz numeroase proteinkinaze, principalele
fiind kinaza lanurilor uoare a miozinei, care induce contracia
muchiului neted i fosforilaz-kinaza, care activeaz fosforilaza i
apoi scindarea glicogenului cu eliberarea glucozei.
Activarea calmodulinei are, printre alte efecte, stimularea
permeabilitii membranei RE pentru Ca
2+
, care revine n lumenul
reticulului endoplasmatic, prin pompa Ca
2+
/ATP-aza.
IP3 poate fi defosforilat de ctre fosfataze specifice cu formare de
IP2 sau fosforilat la IP4 (alt mediator intracelular), compui inactivi.
Ca
2+
- important mesager secund
n celula muscular creterea nivelului citosolic al Ca
2+
declaneaz contracia,
n celulele secretorii declaneaz secreia,
n neuroni declaneaz eliminarea de neurotransmitori
n celula ou iniiaz dezvoltarea embrionului.
activeaz unele enzime
Concentraia Ca
2+
din citoplasm, din RE i mitocondrie se menine la valori sczute
fa de Ca
2+
din lichidele extracelulare (10000 x),
un gradient ce dirijeaz trecerea Ca
2+
prin membran din exterior spre citosol.
Un semnal deschide canalele de Ca
2+
din cele 2 membrane, citoplasmatic si RE
Ca
2+
revine n citoplasm i activeaz proteinele celulare sensibile la Ca
2+
.
DAG rmne la nivelul membranei citoplasmatice unde exercit 2
funcii:
1. poate fi clivat acidul arahidonic eicosanoizii cu rol de
mesageri secundari i hormoni tisulari.
2. activeaz o serin/treonin kinaz, proteinkinaza C (PKC), pe
care o transloc din citoplasm pe faa citoplasmatic a
membranei citoplasmatice. Acolo, PKC este activat de Ca
2+
i
DAG. DAG i Ca
2+
sunt amndoi necesari pentru activarea maxim a
PKC.
PKC activat fosforileaz proteine int care difer funcie de tipul
celulei, fiind implicate n fosforilarea factorilor transcripionali din
nucleu cu rol control n reglarea diviziunii celulare.
Efectele DAG pot fi mimate de esterii forbol, analogi cu DAG, capabili
s activeze direct PKC conducnd la diviziune celular. Diferena
dintre DAG i esterii forbol este aceea c DAG este rapid distrus i
activarea PKC se realizeaz numai cnd este necesar, pe cnd
esterul forbol are via lung i efect protumoral.
Croton tiglium ulei de croton conine forbol
(diterpena) i forbolesteri
GMPc ca mesager secund
GMPc este analog cu AMPc i poate fi obinut din GTP
sub aciunea a 2 guanilat ciclaze (GC1, GC2) diferite
Aceast cale este utilizat de unii hormoni (factor
natriuretic atrial, guanilina, acetilcolina, bradichinina) i
de oxidul nitric.
GMPciclic este un mesager secund care induce efecte
importante n diferite esuturi:
n intestin i rinichi modific transportul ionilor i
reinerea apei.
n muchiul cardiac induce relaxare.
n creier este un element important n procesele
funcionale.
1. Hormoni - se ataeaz la receptorul membranar
specific al crui domeniu intracelular este o guanilat-
ciclaz.
modificare conformaional la nivelul receptorului cu
activarea guanilat ciclazei membranare.
GMPc
GMPc activeaz unele proteinkinaze specifice (PKG).
Hidroliza GMPc de ctre fosfodiesteraze confer
reversibilitate procesului.
Un exemplu este receptorul guanilatciclazei din intestin
care regleaz secreia Cl
n intestin. Receptorul are
ca agonist endotoxina produs de E. coli sau alte
bacterii gram negative. Aciunea acesteia menine
constant un nivel ridicat al secreiei de Cl
fapt ce
reduce reabsorbia apei de ctre intestin, producnd
diaree.
2. Oxidul nitric - rol n comunicarea intercelular n sistemele nervos, imun i
circulator.
exist o a 2-a guanilat ciclaz complet diferit, prezent n citoplasm.
NO este produs la nivelul endoteliului vascular de ctre NO-sintetaza, activat
nervos (acetilcolina).
Arginin + NADPH,H
+
------> citrulin + NO + NADP
+
NO difuzeaz rapid n celulele musculare netede din peretele vascular unde se
leag la fierul din situsul activ al guanilat ciclazei citoplasmatice
GMPc
relaxarea musculaturii netede din peretele vascular i vasodilataia
Alte efecte ale NO
antiagregant plachetar
mediaz aciunea antibacterian i antitumoral a macrofagelor
neurotransmitor n creier
Efecte similare medicamente - nitroglicerina i viagra
Nitroglicerina (tratamentul anginei pectorale) NO
relaxeaz coronarele i reduce sarcina muchiului cardiac.
Viagra (sildenafil) inhib fosfodiesteraza care distruge GMPc
poteneaz efectul NO, a crui producie este crescut de stimularea sexual.
Vasodilataia vaselor de snge peniene menin penisul n erecie.
Aciunea NO pe musculatura neted
Receptori transmembranari de tip
canalele ionice dependente
Excitabilitatea celulelor senzoriale, neuronilor i a
miocitelor este dependent de prezena canalelor ionice
la nivelul crora are loc convertirea semnalelor care
dirijeaz transportul transmembranar al unor ioni
anorganici, precum Na
+
, K
+
, Ca
2+
i Cl
-
, ca rspuns la o
mare varietate de stimuli.
Aceste canale ionice transmembranare se numesc
dependente deoarece pot fi nchise sau deschise n
funcie de activarea receptorului asociat prin legarea
substratului su specific:
un neurotransmitor
modificri ale potenialului electric transmembranar (Vm).
1. Receptorul nicotinic colinergic este un
canal ionic ligand-dependent
Canalul receptorului nicotinic colinergic se deschide ca rspuns la
aciunea neurotransmitorului acetilcolin (i al nicotinei). Acest receptor
se gsete n membrana postsinaptic a neuronilor de la nivelul anumitor
sinapse i n fibrele musculare (miocite) de la nivelul jonciunilor
neuromusculare.
Acetilcolina eliberat de ctre un neuron depolarizat interacioneaz
cu receptorul pentru acetilcolin i determin modificarea conformaiei
receptorului din deschis n nchis.
Influxul de Na
+
sau Ca
2+
va depolariza membrane plasmatic, iniiind
efecte specifice care variaz n funcie de tipul de esut:
potenialul de aciune la nivelul neuronului postsinaptic
depolarizarea fibrei musculare la nivelul jonciunii neuromusculare cu
declanarea consecutiv a contraciei musculare.
Concentraia acetilcolinei la nivelul fantei sinaptice va fi redus rapid
de ctre enzima acetilcolinesteraz, prezent la nivelul sinapsei.
Cnd nivelul acetilcolinei rmne crescut pentru mai mult de cteva
milisecunde, receptorul este desensibilizat, canalul ionic este nchis i
acetilcolina este puternic legat.
2. Neuronii au canale receptor care rspund la aciunea
diferiilor neurotransmitori
Neurotransmitorii, cum ar fi 5-hidroxitriptamina (serotonina),
glutamatul i glicina i pot exercita aciunea prin intermediul
canalelor receptor care au o structur asemntoare cu cea a
receptorului acetilcolinei.
Canalele cationice conin resturi de Glu i Asp, ncrcate
negativ.
n cazul n care aceti aminoacizi acizi sunt nlocuii cu
aminoacizi bazici, canalul cationic este transformat n canal anionic.
n funcie de ionul care trece printr-un canal, ligandul
(neurotransmitorul) pentru acel canal fie depolarizeaz fie
hiperpolarizeaz celula int.
Iniial, membrana plasmatic a neuronului presinaptic este
polarizat (ncrcat negativ n interior) prin aciunea ATP-azei
Na
+
K
+
care determin efluxul a 3 Na
+
consecutiv influxului a 2 K
+
.
(1) Un stimul care acioneaz asupra neuronului determin declanarea
potenialului de aciune care se deplaseaz de-a lungul axonului (sgeata
alb). Deschiderea unui canal de Na
+
voltaj-dependent permite influxul de
Na
+
, iar depolarizarea local astfel rezultat determin deschiderea
canalelor de Na
+
adiacente. Procesul se repet n mod similar. Direcia
deplasrii potenialului de aciune este meninut de o perioad refractar
scurt care se instaleaz n urma deschiderii fiecrui canal de Na
+
voltaj-
dependent.
(2) Cnd unda de depolarizare ajunge la captul distal al axonului,
canalele de Ca
2+
voltaj-dependente se deschid, permind influxul de Ca
2+
n poriunea presinaptic a neuronului.
(3) Concentraia intraneuronal crescut de Ca
2+
favorizeaz
eliberarea, prin exocitoz, a neurotransmitorului acetilcolin n fanta
sinaptic.
(4) Acetilcolina se leag de un receptor de la nivelul neuronului
postsinaptic, stimulnd astfel deschiderea canalului ionic ligand-dependent.
(5) Ionii de Na
+
i Ca
2+
extracelulari intr prin acest canal,
depolariznd neuronul postsinaptic. Astfel, semnalul electric a ptruns n
corpul neuronului postsinaptic i se va deplasa de-a lungul axonului
acestuia, prin aceleai mecanisme, spre un al treilea neuron.
Recepia fascicolului luminos la
nivelul ochiului vertebratelor
Vederea proces dependent de un receptor
cuplat cu o protein G
Fascicolul luminos care ptrunde prin pupil este dirijat ctre retin
care reprezint o reea de neuroni nalt specializai pentru
recepionarea i prelucrarea semnalelor luminoase.
Neuronii fotosenzoriali primari (celulele receptoare) sunt reprezentai
de:
celulele cu bastona (aprox. 10
9
/retin), responsabile de
vederea nocturn (scotopic), dar care nu ofer detalii ale
obiectelor i nu sunt implicate n procesul de difereniere a
culorilor
celulele cu conuri (aprox. 3 X 10
6
/retin) care sunt mai puin
sensibile la lumin, dar sunt responsabile de procesul de
discriminare a culorilor (vederea diurn = fotopic).
Exist 3 tipuri de celule cu conuri. n funcie de pigmenii pe care i
conin, acestea sunt responsabile de recepia celor 3 culori
fundamentale: rou, verde i albastru. Lipsa unuia dintre pigmeni
genereaz discromatopsii, cea mai cunoscut fiind daltonismul.
Att celulele cu conuri ct i cele cu bastona sunt
formate din 2 componente:
segmentul extern care conine un numr mare de discuri
membranoase ncrcate cu proteina membranar, rodopsin,
segmentul intern care conine nucleul i mitocondriile care
genereaz moleculele de ATP necesare n procesul de
fotoconversie.
Celulele cu bastona i cele cu conuri formeaz sinapse
cu o reea de neuroni intermediari (celule bipolare) care
transmit informaia referitoare la activitatea electric
ctre neuronii ganglionari (celule multipolare) localizai n
profunzimea retinei. Neuronii ganglionari integreaz i
transmit influxurile nervoase prin nervul optic ctre
cortexul occipital.
Celula cu bastona (fotoreceptorul cu prelungire n
form de bastona) are 3 seciuni:
partea central care conine nucleul, mitocondriile,
etc.
un capt al celulei care realizeaz o sinaps cu o
celul bipolar conectat la nervul optic
cellalt capt - segmentul extern - sub forma unui
bastona cilindric, coninnd aparatul fototransduciei
: > 2000 de discuri suprapuse. Fiecare disc conine
15 000 molecule de rodopsin (fotoreceptorul).
Rodopsina are legat 11-cis retinal, capabil s
absoarb fotonul
Proteina G asociat este transducina (T
F
) care are
legat GDP
Membrana plasmatic care nconjoar segmentul
extern conine canale de Na
+
, Ca
2+
GMPc
dependente.
Mecanismele declanate ca urmare
a absorbiei de fotoni mediat de
rodopsin la nivelul segmentului
extern al celulei cu bastona
Receptorul luminos din discuri este rodopsina - proteina opsin i 11-
cis-retinal.
Ca i ali receptori asociai proteinei G are 7 helixuri transmembranare.
Cnd rodopsina absoarbe un foton (molecula semnal), are loc o modificare
conformaional n structura acesteia prin transformarea lui 11-cis-retinal
n trans-retinal. Astfel molecula de rodopsin devine activat.
Rodopsina activat determin modificri n alctuirea transducinei, o
protein G trimeric, care schimb GDP n GTP i detaarea subunitii -
GTP.
-GTP migreaz la nivelul membranei citoplasmatice unde activeaz o
fosfodiesteraz, GMPc-fosfodiesteraza, care va hidroliza GMPc.
scade nivelul GMPc din citosol care va determina o scdere a cantitii
de GMPc legat la canalele de Na
+
,Ca
2+
din membrana plasmatic a
segmentului extern al celulei cu bastona.
Canalele de Na
+
se nchid, nceteaz afluxul de Na
+
i Ca
2+
iar potenialul
membranar crete la 70mV.
Astfel semnalul luminos este transformat n semnal electric cu formarea
unui impuls nervos ce strbate nervul optic pn la centrul vederii din SNC.
subunitateai -GTP este inactivat prin activitatea sa GTP-azic.
Transducina revine astfel la stadiul inactiv
Scderea [Ca
2+
] determin activarea guanilciclazei (GC) i inhibarea
PDE; [GMPc] crete depind nivelul de ntuneric, cu redeschiderea
consecutiv a canalelor cationice i revenirea Vm la valoarea
anterioar stimulrii.
Rodopsin kinaza (RK) fosforileaz rodopsina; nivelul sczut al [Ca
2+
] i
recoverinei (Recov) stimuleaz aceast reacie.
Arestina (Ar) inactiveaz rodopsina prin legarea de captul carboxi-
terminal fosforilat al acesteia.
Arestina se disociaz, rodopsina este defosforilat, i trans-retinal
este nlocuit cu 11-cis-retinal. Astfel, rodopsina este pregtit pentru un
nou ciclu de fotoconversie
Transmiterea gustului i mirosului
Are loc similar cu cel al transmiterii semnalului luminos.
Receptorii sunt proteine de tipul receptorilor membranari
cu 7 elici,
Receptorii transmit semnale prin intermediul unor
proteine trimerice de tip G la nivelul adenilatciclazei ce
genereaz AMPc.
G olfactiv pentru miros
gustducin pentru gust
AMPc va deschide canalele ionice depolariznd celula.
Modificri ale cilor de semnalizare n cancer
Genele ce codific proteine implicate n procesul de cretere i diviziune
celular pot fi considerate protooncogene.
Aceste proteine pot fi diferite elemente ale cilor de semnalizare: factori de
cretere, receptori proteine G, protein kinaze sau factori de transcripie
implicai n progresia ciclului celular.
Sub aciunea diferiilor factori: ageni virali, ageni mutageni chimici sau
fizici, rearanjri cromozomiale, protooncogenele sufer mutaii n urma
crora se pot transforma n oncogene. Produsul oncogenei, proteina,
prezint modificri care altereaz funcia normal, amplificnd sau
meninnd permanent activ calea de semnalizare implicat n creterea
sau diviziunea celular.
De exemplu, n cancerul de sn, mutaia genei ErbB, ce codific receptorul
pentru factorul de cretere epidermal (EGFR) produce o form mutant a
receptorului ce menine activ tirozin kinaza indiferent de prezena EGF.
Astfel calea de semnalizare specific factorului de cretere genereaz
continuu semnal de diviziune celular.
Un alt exemplu este al mutaiei genei ce codific proteina Ras n urma
creia aceasta pierde capacitatea GTP-azic. ca urmare Ras GTP
rmne permanent activat genernd un semnal permanent.
Mutaiile proteinelor Ras se ntlnesc n 30 50% din cancerele de
plmn i peste 90% n cele de pancreas.
S-ar putea să vă placă și
- Subiecte Biochimie 2020Document20 paginiSubiecte Biochimie 2020Anonymous 67EwtOk100% (3)
- Subiecte ImunologieDocument116 paginiSubiecte ImunologieDiana IlieÎncă nu există evaluări
- Histologie AN I Sem I AMGDocument10 paginiHistologie AN I Sem I AMGNewLo0k100% (1)
- Reglarea Secretiei Hormonilor TiroidieniDocument16 paginiReglarea Secretiei Hormonilor TiroidieniCristiana IliescuÎncă nu există evaluări
- Numararea Eritrocitelor, Leucocitelor Si TrombocitelorDocument27 paginiNumararea Eritrocitelor, Leucocitelor Si TrombocitelorDiana Cristiana GheorgheÎncă nu există evaluări
- Leucocite CURSDocument71 paginiLeucocite CURSNico IonaşcuÎncă nu există evaluări
- Subiecte Examen Fiziologie MG I SEM II SibiuDocument4 paginiSubiecte Examen Fiziologie MG I SEM II Sibiumaria_fratila_2Încă nu există evaluări
- Biochimia Este Ştiinţa Care Studiază Bazele Moleculare Ale VieţiiDocument481 paginiBiochimia Este Ştiinţa Care Studiază Bazele Moleculare Ale VieţiivaliÎncă nu există evaluări
- 4 Embriopatii FetopatiiDocument8 pagini4 Embriopatii FetopatiiRaluka UkaÎncă nu există evaluări
- Fiziopatologia SoculuiDocument10 paginiFiziopatologia SoculuiOvidiu Aioanei100% (1)
- Explorarea Proteinelor PlasmaticeDocument9 paginiExplorarea Proteinelor PlasmaticeAlex SeiceanÎncă nu există evaluări
- Electroforeza in Gel de AgarozaDocument5 paginiElectroforeza in Gel de Agarozamanu_cyb100% (1)
- Subiecte Biologie Celulara UMF Carol DavilaDocument2 paginiSubiecte Biologie Celulara UMF Carol DavilaAlexandra MarinÎncă nu există evaluări
- TriglicerideDocument15 paginiTriglicerideconstantin67% (3)
- Insulina GlucagonDocument44 paginiInsulina GlucagonAlexandru IarovoiÎncă nu există evaluări
- DETERMINAREA AMILAZEI CapatanDocument19 paginiDETERMINAREA AMILAZEI CapatanKis Marta TamaraÎncă nu există evaluări
- ReceptoriiDocument96 paginiReceptoriiAndreea IvanescuÎncă nu există evaluări
- Curs 3 Aparatul GolgiDocument4 paginiCurs 3 Aparatul GolgiOana Koppandi100% (1)
- Biochimie 7 - Metabolismul Glucidelor I (Glicoliza, Decarboxilarea, Ciclul Krebs) CATB (An III)Document55 paginiBiochimie 7 - Metabolismul Glucidelor I (Glicoliza, Decarboxilarea, Ciclul Krebs) CATB (An III)Alex Andru100% (2)
- HEMOPROTEINEDocument6 paginiHEMOPROTEINEGherman MadalinaÎncă nu există evaluări
- GlicolizaDocument13 paginiGlicolizaAndreea Ciomaga100% (2)
- 07 Aplicatii Practice in Lab de BiochimieDocument39 pagini07 Aplicatii Practice in Lab de BiochimieEnache CristinaÎncă nu există evaluări
- Metabolismul ProteinelorDocument12 paginiMetabolismul ProteinelorCalin-Sorin CimpeanuÎncă nu există evaluări
- Cuers 1 Biochimie Anul I MedicinaDocument23 paginiCuers 1 Biochimie Anul I MedicinaAlexandru BarbuÎncă nu există evaluări
- Dogma CentralaDocument60 paginiDogma CentralaLoghin Andrei50% (2)
- Indreptar de Lucrari Practice PDFDocument123 paginiIndreptar de Lucrari Practice PDFHedes Despina100% (1)
- 1b Lanțul Transportor de ElectroniDocument82 pagini1b Lanțul Transportor de ElectroniLaura HutzelÎncă nu există evaluări
- Metabolismul GlucidicDocument48 paginiMetabolismul Glucidicanddrutza100% (1)
- Calea Pentozofosfat - Calea UronicaDocument12 paginiCalea Pentozofosfat - Calea UronicaAlex DanciuÎncă nu există evaluări
- Hormoni PDFDocument45 paginiHormoni PDFlilip6Încă nu există evaluări
- Ischemie Leziune NecrozaDocument83 paginiIschemie Leziune NecrozaVisan Andreea Mihaela Ramona100% (1)
- FiziopatologieDocument30 paginiFiziopatologieanne9oÎncă nu există evaluări
- Lp-Fiziologie An I 2012Document108 paginiLp-Fiziologie An I 2012aaancutzaaaÎncă nu există evaluări
- 1+2. Acizii Nucleici Si Functiile Acizilor NucleiciDocument60 pagini1+2. Acizii Nucleici Si Functiile Acizilor Nucleicicarmen_derscariu3738100% (1)
- Lipoproteinele PlasmaticeDocument19 paginiLipoproteinele PlasmaticeMihaela CarnutaÎncă nu există evaluări
- Explorarea Metabolismului ProtidicDocument6 paginiExplorarea Metabolismului ProtidicRusu Stefan100% (1)
- Biologie Celulara - Adeziunea CelularaDocument9 paginiBiologie Celulara - Adeziunea CelularaLaura Pana100% (1)
- Glicoliza. GluconeogenezaDocument12 paginiGlicoliza. GluconeogenezaLaura Cristina100% (2)
- Curs Enzime - Biochimie Anul IDocument74 paginiCurs Enzime - Biochimie Anul IFlavius CristuineaÎncă nu există evaluări
- L 4 - Dozarea Proteinelor Prin Metoda LowryDocument2 paginiL 4 - Dozarea Proteinelor Prin Metoda LowryRoxana RoxyÎncă nu există evaluări
- 2016 - Taus - Teste de Toxicologie ClinicaDocument229 pagini2016 - Taus - Teste de Toxicologie ClinicaAnonymous IOrWuyVQS1Încă nu există evaluări
- OperonulDocument20 paginiOperonulAna-Maria LopotencoÎncă nu există evaluări
- AQUAPORINEDocument26 paginiAQUAPORINEProca IonÎncă nu există evaluări
- Curs Biochimie Sem IIDocument124 paginiCurs Biochimie Sem IIbucureanionut100% (3)
- Amprentarea GenomicaDocument21 paginiAmprentarea GenomicaAlinutza PowerpuffÎncă nu există evaluări
- Numararea HematiilorDocument28 paginiNumararea HematiilorCristina Criss100% (2)
- Fiziologie (Cursul 5)Document27 paginiFiziologie (Cursul 5)laviniapatrascoiuÎncă nu există evaluări
- Transmiterea SinapticaDocument16 paginiTransmiterea SinapticaMădălina NăstuţăÎncă nu există evaluări
- Subiecte BiochimieDocument43 paginiSubiecte BiochimieBarbuta Cristina Elena100% (1)
- Metabolismul AminoacizilorDocument38 paginiMetabolismul AminoacizilorMitrea Andreea Cristina100% (3)
- Grile Examen BiocelDocument178 paginiGrile Examen BiocelTeodora DumitrașcuÎncă nu există evaluări
- Gluconeogeneza 2022 PDFDocument11 paginiGluconeogeneza 2022 PDFLaura MunteanuÎncă nu există evaluări
- TolerantaDocument10 paginiTolerantaBianca AndreeaÎncă nu există evaluări
- Hormonii Implicati in Mentinerea Echilibrului Fosfo Cu DiacriticeDocument9 paginiHormonii Implicati in Mentinerea Echilibrului Fosfo Cu Diacriticeniko55555Încă nu există evaluări
- C12-13semnalizarea CelularăDocument35 paginiC12-13semnalizarea CelularăCezara AxinteÎncă nu există evaluări
- Semnalizarea CelularaDocument71 paginiSemnalizarea CelularaRusu CristianaÎncă nu există evaluări
- CURS2 BiocelDocument131 paginiCURS2 Biocelexplorediscover100% (1)
- Icurs10 11 1Document70 paginiIcurs10 11 1maÎncă nu există evaluări
- Aspecte Generale Ale Semnalizării CelulareDocument65 paginiAspecte Generale Ale Semnalizării CelulareAlina MihuÎncă nu există evaluări
- 4 Semnale Inter - Si Intra-CelulareDocument70 pagini4 Semnale Inter - Si Intra-CelulareCostin AleodorÎncă nu există evaluări
- Metabolism Acizi NucleiciDocument94 paginiMetabolism Acizi NucleiciOvidiu VaidaÎncă nu există evaluări
- Testele Screening, de Inteligenta, de AptitudiniDocument25 paginiTestele Screening, de Inteligenta, de AptitudiniOvidiu VaidaÎncă nu există evaluări
- Sinteza ProteinelorDocument35 paginiSinteza ProteinelorOvidiu VaidaÎncă nu există evaluări
- Arnica MontanaDocument3 paginiArnica MontanaOvidiu VaidaÎncă nu există evaluări