S-ar putea să vă placă și
- BiofizicaDocument6 paginiBiofizicaMohammed ArarÎncă nu există evaluări
- SondajeDocument27 paginiSondajeAndreia PaladeÎncă nu există evaluări
- 01.structura MicroscopicaDocument3 pagini01.structura MicroscopicaGabi VasiliuÎncă nu există evaluări
- Domeniile Biofizicii 01.03.2021Document3 paginiDomeniile Biofizicii 01.03.2021Mirela IleanaÎncă nu există evaluări
- ReferatDocument7 paginiReferatCarinaDianaÎncă nu există evaluări
- 39.seroterapia Si VaccinoterapiaDocument2 pagini39.seroterapia Si VaccinoterapiaIasmina MihaelaÎncă nu există evaluări
- Decubit LateralDocument1 paginăDecubit LateralIonut DinamovistulÎncă nu există evaluări
- Curs 1 Importanta Studiului Anatomiei Umane Pentru AmgDocument13 paginiCurs 1 Importanta Studiului Anatomiei Umane Pentru Amgalfa5017Încă nu există evaluări
- TestDocument3 paginiTestDaniela LutacÎncă nu există evaluări
- 2 Nevoia de A RESPIRA Și A AVEA O BUNĂ CIRCULAȚIEDocument23 pagini2 Nevoia de A RESPIRA Și A AVEA O BUNĂ CIRCULAȚIEDumi DumiiÎncă nu există evaluări
- Flora NormalaDocument2 paginiFlora NormalaBratescuCristian100% (1)
- Ultrasunetele ReferatDocument6 paginiUltrasunetele ReferatgeorgyÎncă nu există evaluări
- Biofizica Si BiomecanicaDocument6 paginiBiofizica Si BiomecanicaCosmin Valentin100% (1)
- Migratia Si SegmentatiaDocument3 paginiMigratia Si SegmentatiaIulia Vasilica GirosanÎncă nu există evaluări
- Excitabilitatea - BiofizicaDocument21 paginiExcitabilitatea - Biofizicamarius_inter881956Încă nu există evaluări
- Asepsie & AntisepsieDocument8 paginiAsepsie & AntisepsieGabriela Ximena CaraivanÎncă nu există evaluări
- Placenta ReferatDocument9 paginiPlacenta Referatanon_298347676Încă nu există evaluări
- Transportul Bolnavului Cu Fotoliu RulantDocument2 paginiTransportul Bolnavului Cu Fotoliu RulantinaÎncă nu există evaluări
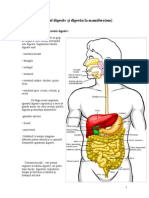
- Aparatul Digestiv Şi Digestia La MamifereDocument4 paginiAparatul Digestiv Şi Digestia La MamifereDiana AdamciucÎncă nu există evaluări
- Instilatie OcularaDocument2 paginiInstilatie Ocularap_983820008Încă nu există evaluări
- Fisa 1 - Administrarea Antibioticelor Pe Cale Parenteral ÂDocument1 paginăFisa 1 - Administrarea Antibioticelor Pe Cale Parenteral ÂOana Diana AeleneiÎncă nu există evaluări
- 1 - Nevoia de A ComunicaDocument4 pagini1 - Nevoia de A ComunicaJustyLynAbyÎncă nu există evaluări
- Hipnotice PDFDocument20 paginiHipnotice PDFconstantinÎncă nu există evaluări
- Perioada EmbrionaraDocument2 paginiPerioada EmbrionaraReihan Sadîk100% (1)
- Administrarea Medicamentelor Pe Cale PercutanatăDocument4 paginiAdministrarea Medicamentelor Pe Cale PercutanatăRaluca LunguÎncă nu există evaluări
- Tesutul ConjunctivDocument3 paginiTesutul ConjunctivandreeaÎncă nu există evaluări
- Nevoia de A Evita PericoleleDocument3 paginiNevoia de A Evita PericoleleAdelaida NegulescuÎncă nu există evaluări
- Fiinta Umana Si Nursing-Ul - Nevoia 8Document3 paginiFiinta Umana Si Nursing-Ul - Nevoia 8Violeta Dragomir100% (1)
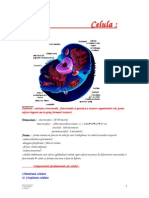
- CelulaDocument7 paginiCelulagoroonÎncă nu există evaluări
- Autoexaminarea SanilorDocument12 paginiAutoexaminarea SanilorpaulappopaÎncă nu există evaluări
- ProteazomiDocument80 paginiProteazomiLuiza Musat100% (2)
- Coci PatogeniDocument24 paginiCoci PatogeniMinciuna AlinaÎncă nu există evaluări
- Referat Igiena Cojocariu Denisa Iuliana Boli-Transmise-Prin-ApăDocument7 paginiReferat Igiena Cojocariu Denisa Iuliana Boli-Transmise-Prin-ApăCristea GabrielÎncă nu există evaluări
- Dezvoltarea Aparatului ReproducaorDocument2 paginiDezvoltarea Aparatului ReproducaorAndreea VieruÎncă nu există evaluări
- Curs Digestie 1Document38 paginiCurs Digestie 1Ioana Oprea100% (1)
- MICROBIOLOGIEDocument39 paginiMICROBIOLOGIEaxiel90100% (1)
- Caile Si Mecanismele de Transmitere Ale Agentilor InfectiosiDocument12 paginiCaile Si Mecanismele de Transmitere Ale Agentilor InfectiosiAna Maria Stan100% (1)
- Echinococoza Si BotriocefalozaDocument19 paginiEchinococoza Si BotriocefalozaRadu Leca100% (1)
- Curs 3 SemiologieDocument26 paginiCurs 3 SemiologieMihai Georgian IonitaÎncă nu există evaluări
- Calitatile PulsuluiDocument1 paginăCalitatile PulsuluiTamara CristureanuÎncă nu există evaluări
- Curs Biofizica CelulaDocument13 paginiCurs Biofizica CelulaIzabellaÎncă nu există evaluări
- Nursingul Pacientului Cu ConstipatieDocument94 paginiNursingul Pacientului Cu ConstipatieRobert StratÎncă nu există evaluări
- Maduva SpinariiDocument4 paginiMaduva SpinariitudortudorgÎncă nu există evaluări
- Curs Amg Singele Si T.muscularDocument53 paginiCurs Amg Singele Si T.muscularAdina VioÎncă nu există evaluări
- ChlamidiiDocument61 paginiChlamidiilacatisÎncă nu există evaluări
- Educatie Pentru Sanatate t1-2Document3 paginiEducatie Pentru Sanatate t1-2free_loÎncă nu există evaluări
- Recapitulare - Componentele Materiei Vii, MetabolismulDocument17 paginiRecapitulare - Componentele Materiei Vii, Metabolismuliulia100% (1)
- AnalizatoriiDocument20 paginiAnalizatoriiRedEyes09Încă nu există evaluări
- Informatii Care Te Ajuta Sa Descifrezi Un Buletin de Analize MedicaleDocument2 paginiInformatii Care Te Ajuta Sa Descifrezi Un Buletin de Analize MedicaleIuliaAdrianaÎncă nu există evaluări
- Administrarea MedicamentelorDocument52 paginiAdministrarea Medicamentelormihai bibiÎncă nu există evaluări
- 2 Structura Si Functia CeluleiDocument4 pagini2 Structura Si Functia CeluleiEcaterina BabanutaÎncă nu există evaluări
- Curs BiofizicaDocument13 paginiCurs BiofizicaIla MariusÎncă nu există evaluări
- Dezobstruarea Cailor RespDocument16 paginiDezobstruarea Cailor RespDani DanielaÎncă nu există evaluări
- Membrane Biologice 1Document10 paginiMembrane Biologice 1Anca NegreanÎncă nu există evaluări
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Fiziologie Tot1Document18 paginiFiziologie Tot1Maria Moiseencu100% (1)
- Membrane Biologice - BunDocument21 paginiMembrane Biologice - BunMarius Alexa67% (3)
- Curs 4 BIOFIZDocument72 paginiCurs 4 BIOFIZfloricapopovici168Încă nu există evaluări
- Biofizica LPDocument12 paginiBiofizica LPIulya BraghesÎncă nu există evaluări
- Transportul Prin Membrana CelularaDocument7 paginiTransportul Prin Membrana CelularaTeodora Elena100% (1)
- Grile Licenta IasiDocument196 paginiGrile Licenta Iasijollie_elleÎncă nu există evaluări
- Insuficienta RespiratorieDocument14 paginiInsuficienta RespiratoriePetrisor VulpeÎncă nu există evaluări
- Numarul de Aur in StomatologieDocument1 paginăNumarul de Aur in StomatologieKeyser SozeÎncă nu există evaluări
- Insuficienta Respiratorie AcutaDocument14 paginiInsuficienta Respiratorie AcutaPetrisor Vulpe100% (1)
- Insuficienta Respiratorie AcutaDocument14 paginiInsuficienta Respiratorie AcutaPetrisor Vulpe100% (1)
- De Foarte Mult Timp Am Dorit Sa Fac Un Asemenea ArticolDocument28 paginiDe Foarte Mult Timp Am Dorit Sa Fac Un Asemenea ArticolPetrisor Vulpe100% (1)
- Bazinul OsosDocument6 paginiBazinul OsosPetrisor VulpeÎncă nu există evaluări
- Canalul Moale ObstetricalDocument15 paginiCanalul Moale ObstetricalPetrisor Vulpe100% (1)
- Circulatia Apei in Organismul Uman: Universitatea Apollonia Din IasiDocument9 paginiCirculatia Apei in Organismul Uman: Universitatea Apollonia Din IasiPetrisor VulpeÎncă nu există evaluări
- Durerea DentinaraDocument20 paginiDurerea Dentinaramikitoma100% (1)
- Durerea DentinaraDocument20 paginiDurerea Dentinaramikitoma100% (1)
- Anatomie II LP 1Document15 paginiAnatomie II LP 1Petrisor VulpeÎncă nu există evaluări
- GEnetecaDocument21 paginiGEnetecaPetrisor VulpeÎncă nu există evaluări
- Fiziologie Curs 1Document3 paginiFiziologie Curs 1Petrisor VulpeÎncă nu există evaluări
- ReferatDocument7 paginiReferatPetrisor VulpeÎncă nu există evaluări