CAPITULO 2
TaBia 2-1. Calores de
vaporizacién de algunos I
corrientes en sus puntos de
ebullicién (1,0 atm).
Liquido Ap
cal
Agua 540
Metanol 263
Etanol 204
n-Propanol 164
Acetona 125
Benceno 94
Cloroformo 59
El agua
El agua no solamente constituye del 70 al 90 por ciento del
peso de la mayor parte de las formas vivas, sino que repre-
senta la fase continua de los organismos. A causa de su
abundancia y ubicuidad el agua es considerada, con frecuen-
cia, un liquido inerte, meramente destinado a Ienar espacios
en los organismos vivos. Pero, en realidad, el agua es una
sustancia de gran reaccionabilidad, con propiedades poco fre-
cuentes, que la diferencian mucho, tanto fisica como quimica-
mente, de la mayoria de los liquidos corrientes. Sabemos ahora
que el agua y los productos de su ionizacién, los iones hidr6-
geno e hidroxilo, son factores importantes en la determinacién
de la estructura y las propiedades bioldgicas de las proteinas,
de los Acidos nucleicos, asi como de las membranas, de los
ribosomas y de otros muchos componentes celulares.
Propiedades fisicas y enlace de hidrégeno en el agua
El agua posee un punto de fusién, un punto de ebullicién, el
calor de vaporizacién y el de fusién y la tensién superficial,
mas elevados que otros hidruros comparables, tales como el
HS 0 el NH, o para el caso, que la mayor parte de los
Tiquidos corrientes. Todas estas propiedades indican que en
el agua las fuerzas de atraccién entre las moléculas y, por
tanto, su cohesién interna, son relativamente elevadas. Por
ejemplo, la tabla 2-1 muestra que el calor de vaporizacién
del agua es considerablemente mayor que el de cualquier otro
de los liquidos corrientes de la lista. El calor de vaporizacién
es una medida directa de la cantidad de energia necesaria
para superar las fuerzas de atraccién entre las moléculas adya-
centes en un liquido, de modo que las moléculas individuales
puedan separarse unas de otras y pasar al estado gaseoso.
Las potentes fuerzas intermoleculares en el agua liquida
estan originadas por la distribucién especifica de los electro-
nes en la molécula de agua (fig. 2-1). El atomo de oxigeno
comparte un par de electrones con cada uno de los atomos
de hidrégeno, por superposicién de los orbitales 1s de los
atomos de hidrégeno con los orbitales hibridos sp? del atomo
de oxigeno. Mediante anilisis espectroscépico y de rayos X
se ha determinado el angulo de enlace H—O—H, que es de
104,5° y la distancia interatémica media H—O que es
41
PARTE 1 COMPONENTES MOLECULARES DE LAS CALULAS
0,0965 nm (fig. 2-1). Esta disposicién de electrones en la
molécula del agua le comunica asimetria eléctrica. E] atomo de
oxigeno, mas electronegativo, tiende a atraer los electrones
no compartidos del atomo de hidrdgeno, y deja desnudos los
nicleos de hidrégeno. El resultado es que cada uno de los dos
atomos de hidrégeno posee una carga local parcial positiva
(designada por 8°). El tomo de oxigeno, a su vez, posee una
carga local parcial negativa (designada por 8-), situada en
la zona de los orbitales no compartidos. Asi, aunque la molé-
cula de agua no posee una carga neta, es un dipolo eléctrico.
Cuando dos moléculas de agua se aproximan mucho, se
establece una atraccién electrostatica entre la carga parcial
negativa situada sobre el atomo de oxigeno de la molécula
de agua y la carga parcial positiva situada sobre un atomo
de hidrégeno de una molécula de agua adyacente. Ello va
acompafiado por una redistribucién de las cargas electrénicas
en ambas moléculas, lo cual exalta en gran medida su interac-
cién. Una unién electrostatica compleja de esta clase se llama
enlace de hidrégeno, Debido a la ordenacién casi tetraédrica
de los electrones alrededor del atomo de oxigeno, cada molé-
cula de agua es potencialmente capaz de unirse mediante
enlaces de hidrégeno con cuatro moléculas de agua vecinas
(fig. 2-2). Esta propiedad es la responsable de Ia elevada
cohesién interna del agua liquida,
Los enlaces de hidrégeno son relativamente débiles com-
parados con los enlaces covalentes. Se calcula que los enlaces
de hidrégeno en el agua liquida poseen solamente una energia
de enlace de unas 4,5 kcal mol, en comparacién con las
110 kcal mol! de los enlaces,covalentes H—O de la molécu-
la de agua (téngase en cuenta que Ja energia de enlace es la
energia necesaria para romper un enlace). Otra propiedad
importante de los enlaces de hidrégeno es que su fortaleza
Ficura 2-2
(Abajo) Enlace tetraédrico de hidesgeno
alrededor de una molécula de agua en
el hielo. Las moléculas 1 y 2 y la
molécula central se hallan en el plano
del papel: ta molécula 3 se halla
por encima de él, y la molécula 4,
detras del plano. (Derecha) Diagrama
esquemitico del reticulo de motéculas de
agua en el hielo,
m
OS
\
i
pr
8
5
&
3
O-n
42
Fioura 2-1
Molécula de agua. El bosquejo det
modelo espacial representa el confin en
el que las atracciones de van der Waals
estan equilibradas por las fuerzas de
repulsion. Se muestra mas abajo
un modelo cristalografico, o de bolas
y varillas, de la molécula de agua con
el angulo de valencia y la longitud.
Radio de van der
‘Waals del atomo de
hidrégeno = 0,12 nm
Longitud del enlace
covalente = 0,0965 nm
Radio de van der
‘Waals del stomo de
oxigeno = 0,14 nm
Direccién del
momento dipolar
Ficura 2-3
Caricter direccional de, los enlaces de
hidrogeno. (Atriba) Cuando el enlace
O-H es lineal respecto al dfomo aceptor,
el enlace de hidesgeno formado tendra
la maxima estabilidad. (Abajo) Cuando
el oxigeno aceptor no se halla sobre ef
vector eléctrico del enlace O—H, se
forma un enlace de hideégeno mas debit.
Capitulo 2 El agua
es maxima cuando los dos grupos que interaccionan se hallan
orientados de modo que proporcionen el maximo de atraccién
electrostatica (fig. 2-3). También los enlaces de hidrégeno
poseen una longitud de enlace caracteristica, la cual difiere
de un tipo de enlace de hidrégeno a otto, que depende de la
geometria estructural y de la distribucién electronica en las
moléculas unidas. En el hielo, por ejemplo, todas las moléculas
de agua se hallan unidas mediante enlaces de hidrégeno; la
longitud del mencionado enlace es de 0,177 nm (fig. 2-2).
Estructura del agua liquida
El establecimiento de puentes de hidrégeno entre las moléculas
de agua no sucede solamente en estado liquido, sino también
en el hielo y en la fase de vapor. En Ja forma cristalina mas
corriente del hielo, conocida como hielo I, cada molécula de
agua se halla unida mediante enlaces de hidrégeno a exacta-
mente las cuatro moléculas de agua mas proximas, constitu-
yendo una red regular, y la distancia media oxigeno-oxigeno
es de 0,276 nm. En el agua liquida cada molécula de agua
a 0°C se halla unida en cualquier momento dado, por tér-
mino medio, a otras 3,6 moléculas de agua; la distancia medi:
oxigeno-oxigeno es sélo ligeramente mayor que en el hielo,
cerca de 0,29 nm a 15°C y 0,305 nm a 83°C. Se ha calculado,
a partir del calor de fusién del hielo, que cuando éste se
funde para producir agua a 0°C solamente se rompen el
10% de los enlaces de hidrégeno del hielo. Asi el agua li-
quida presenta una ordenacion considerable en un radio pe-
quefio aunque esta estructuraciér no se extiende a gran distan-
cia. El agua liquida esta ain altamente ligada por puentes de
hidrégeno a 100°C, como se desprende de sus altos calor de
vaporizacién y constante dieléctrica.
La pequefia diferencia que existe entre el agua liquida y el
hielo en cuanto a la proporcién de enlaces de hidrégeno puede
parecer sorprendente si se considera la rigidez del hielo y la
fluidez del agua. Parte de la explicacién reside en la muy
elevada velocidad con que se forman y se escinden los enlaces
de hidrégeno en el agua liquida. Aungue en un instante dado
la mayor parte de las moléculas del agua liquida se hallan
unidas mediante enlaces de hidrégeno, la vida media de cada
uno de los enlaces de hidrégeno es solamente de 10-" s. La
estructura de corto radio del agua liquida es, por tanto, esta-
distica, ya que es promediada tanto en el espacio como en el
tiempo. Por consiguiente el agua liquida, a la vez que es fluida,
se halla también altamente dotada de enlaces de hidrégeno.
Se han propuesto muchos modelos para la estructura del
agua liquida, aunque ninguno de ellos ha sido comprobado
experimentalmente. Los modelos mas sencillos sugieren que el
agua liquida esta constituida por agrupaciones de moléculas
de agua semejantes al hielo, en equilibrio 1abil con moléculas
de agua libres, Otros modelos proponen que el agua liquida
contiene tres 0 mas tipos de componentes unidos por enlaces
de hidrégeno. Un nuevo tipo, el modelo continuo, sugiere que
aunque la gran mayoria de los enlaces de hidrégeno existen-
tes entre las moléculas de agua en el hielo a 0°C permanecen
sin romperse cuando se funde el hielo, experimentan, sin em-
bargo, distorsiones; es decir, propenden a diferentes angulos
desde la configuracién lineal mas estable mostrada en la fi-
be:
PARTE 1 COMPONENTES MOLECULARES DE LAS CALULAS
gura 2-3. Cuanto mayor es la temperatura del agua liquida,
mayor es la distorsién y la inestabilidad. En este modelo, las
propiedades individuales de vida corta de las moléculas de
agua, aunque poseen éstas gran proporcién de enlaces de hi-
drégeno, se desvian significativamente de la estructura reti-
cular cristalina del hielo. Esta claro que la prosecucién de las
investigaciones acerca de la estructura del agua liquida sera del
mayor significado para la bioquimica y la biologia molecular.
Otras propiedades de los enlaces de hidrégeno
Los enlaces de hidrégeno no se presentan solamente en el
agua. Tienden a formarse entre cualquier atomo electrone-
gativo como el oxigeno, el nitrégeno o el flior, y un atomo
de hidrégeno unido covalentemente a otro atomo electronega~
tivo. Los enlaces de hidrégeno pueden formarse entre dos
moléculas 0 entre dos partes de una misma molécula. En la
figura 2-4 aparecen algunos ejemplos de enlaces de hidrégeno
biolégicamente importantes,
Cuando dos moléculas estan unidas por un solo enlace de
hidrégeno, el enlace sera muy débil en un sistema acuoso
porque las moléculas de agua circundantes competiran para
formar enlaces de hidrégeno con las moléculas de soluto. No
obstante, cuando resulta posible el establecimiento de dos o
mas enlaces de hidrégeno entre dos moléculas de soluto, por
razones geométricas el establecimiento del primer enlace de
hidrégeno incrementa grandemente la posibilidad de que se
forme el segundo. Una vez se ha establecido éste, se incre-
menta la probabilidad de que se forme un tercero, y asi suce-
sivamente, conduciendo a und asociacién muy fuerte entre las
dos moléculas de soluto, las cuales pueden superar los efectos
de competencia de las moléculas del agua. La intensificacién
de la fuerza de atraccién entre dos moléculas por la coopera-
cién de muchos enlaces débiles se llama cooperatividad.
El enlace de hidrégeno cooperativo es una caracteristica
exhibida tanto por las proteinas (pag. 132) como por los
Acidos nucleicos (pag. 877), cuyas moléculas pueden conte-
ner docenas, centenares y atin millares de enlaces de hidré-
geno cooperativos. La cooperatividad del enlace de hidrégeno
en las macromoléculas biolégicas depende de las posiciones
relativas de los grupos funcionales capaces de formar enlaces
de hidrégeno, Es algo parecido a la construccién de una esca-
lera. Una vez se han colocado uno o dos travesafios en su
lugar, de modo que las guias laterales se hallen alineadas, el
resto de los travesafios puede fijarse rapidamente.
Los enlaces de hidrégeno se forman y se rompen mucho
més rapidamente en los sistemas acuosos que la mayor parte
de los enlaces covalentes. Este hecho, junto con su especifi-
cidad geométrica y su cardcter direccional, proporciona a los
enlaces de hidrégeno una gran ventaja biolégica sobre los
enlaces covalentes en los fenémenos biomoleculares que deben
producirse a velocidades muy elevadas, tales como el plega-
miento de las proteinas en sus conformaciones nativas (pa-
ginas 146 y 147),
Propiedades disolventes del agua
El agua es un disolvente mucho mejor que la mayor parte
de los liquidos corrientes. Muchas sales cristalizadas y otros
44
Figura 2-4
Algunos enlaces de hidrégeno de
importancia biolégica.
Entre un RL
grupo °
hidroxilo A
y HO
0,
Entre un RL
grupo
carbonilo
y HO
Entre dos R R
cadenas
Entre pares
de bases
complementarias
en el DNA \),-©
Tiina
Adenina
‘Tapia 2-2. Constantes
dieléctricas de algunos liquidos
a 20°C,
Liquido D
Agua 80
‘Metanol 33
Etanol 24
Acetona 214
Benceno 23
Hexano 19
Figura 2-5
Formacién de una micela de jabén en
agua. Las cadenas no polares del oleato
sédico se ocultan al agua, mientras que
os grupos carboxilo, cargados
negativamente, se hallan expuestos.
‘¢
‘a
2
aN,
A
Simbolismo
Nol
=
Cabeza
aA
SAS
ak
Cadena
Cadena|
polar
VY
=
CH,
AB
ICH,
2
ie
g
Oleato sédico
Micela de oleato sédico
Capitulo 2 El agua
compuestos iénicos se disuelven con facilidad en el agua,
mientras son casi insolubles en los liquidos no polares, tales
como el cloroformo o el benceno, Puesto que la red cristalina
de las sales, por ejemplo el cloruro sédico, se mantiene unida
mediante fuertes atracciones electrostaticas entre iones posi-
tivos e iones negativos alternantes, se necesita de una energia
considerable para separar a estos iones unos de otros. El agua
disuelve, no obstante, el cloruro sédico cristalizado, gracias
a las fuertes atracciones electrostaticas entre los dipolos del
agua y los iones Na* y Cl-, que forman los iones hidratados
correspondientes, muy estables, y superan con ello la tenden-
cia de los iones Na* y Cl a atraerse mutuamente.
El agua tiende también a oponerse a la atraccién electros-
tatica entre los iones positivos y negativos. Esta tendencia
viene expresada por la constante dieléctrica, D, definida por
la relacion
en la que F es la fuerza de atraccién entre dos iones de carga
opuesta, ¢; y € son las cargas de los iones y r es la distancia
entre ellos. Como puede verse en la tabla 2-2, el agua posee
una constante dieléctrica relativamente alta y la correspon-
diente al benceno es muy pequefia. Las fuerzas de atraccién
en el agua entre los iones Nat y Cl a una determinada di
tancia, son solamente un catorceavo de las que mostrarian en
el benceno; es un factor que favorece mucho la disolucién
de la red cristalina por el agua.
Una amplia segunda clase de sustancias que se disuelven
en el agua con facilidad, comprende compuestos no iénicos
pero de caracter polar, tales como los azticares, los alcoholes
sencillos, los aldehidos y las cetonas. Su solubilidad se debe
a la tendencia de las moléculas de agua a establecer enlaces
de hidrégeno con grupos funcionales polares, tales como los
grupos hidroxilo de los azicares y de los alcoholes y el atomo
de oxigeno del grupo carbonilo de los aldehidos y de las
cetonas (fig. 2-4)
Interacciones hidrofébicas
El agua también dispersa 0 solubiliza formando micelas a
muchos compuestos que contienen grupos simultaneos, fuerte
mente no polares y grupos fuertemente polares. Tales molé-
culas reciben el nombre de anfipaticas.
Un ejemplo sencillo de biomolécula anfipatica que tiende
a formar micelas es la sal s6dica del acido oleico, acido graso
de cadena larga, Esta molécula posee un solo grupo carboxilo,
que es polar y tiende por ello a hidratarse con facilidad, y
una larga cola hidrocarbonada, que es no polar e intrinseca-
mente insoluble en el agua. Debido a su larga cola hidrofébica,
el oleato sédico (un jabén) tiene muy poca tendencia a disol-
verse en el agua en forma de una disolucién molecular verda-
dera. Se dispersa en ella con facilidad, sin embargo, con
formacién de micelas en las que los grupos carboxilo negati-
vamente cargados se hallan expuestos a la fase acuosa y los
grupos no polares permanecen ocultos dentro de la estruc-
tura micelar (fig. 2-5). Las micelas de jabén poseen una carga
negativa neta y permanecen en suspensién debido a sus mu-
45
PARTE 1 COMPONENTES MOLECULARES DE LAS CELULAS
tuas repulsiones de naturaleza electrostatica. Tales micelas,
que pueden contener centenares y aun millares de moléculas
de jabén, se forman espontaneamente ya que el agua «pre-
fiere> al agua (y también a los grupos carboxilo) mas que a
las estructuras no polares. En el interior de las micelas las
interacciones de Van der Waals’ aportan fuerzas de atrac-
cién adicionales entre las estructuras hidrocarbonadas adya-
centes,
Debe resaltarse que no existe un enlace estequiométrico
verdadero entre los grupos hidrocarbonados de una micela.
Por dicha razén resulta mas apropiado emplear el término
interaccién hidrofobica que el de «enlace hidrofobico», para re-
Teritse a Ta asociacién de las porciones hidrofébicas de las
moléculas anfipaticas. En comparacién con los enlaces de
hidrégeno, las interacciones hidrofbicas poseen poco caracter
direccional, pero tienden a producir sistemas de elevada esta-
bilidad.
Como veremos mas adelante, otros muchos componentes
celulares son también anfipaticos y tienden a formar estruc-
turas en las que las partes no polares, hidrofébicas, se hallan
ocultas al agua, especialmente en las proteinas y en los acidos
nucleicos (pag. 145) y en los lipidos polares (pag. 294).
Efecto de los solutos sobre las propiedades del agua
La presencia de solutos disueltos provoca cambios en la es-
tructura y en las propiedades del agua liquida. Por ejemplo,
cuando una sal tal como NaCl se disuelve en el agua, los
iones Na’ y Cl, se hallan redeados de una capa de dipolo
del agua. Estos iones hidratados poseen una geometria algo
diferente de las agrupaciones de moléculas de agua unidas
mediante enlaces de hidrégeno; su ordenacién superior y de
estructura es mas regular. La tabla 2-3 muestra las distancias
interiénicas medias en disoluciones acuosas de NaCl en fun-
cién de la concentracién de Ia sal. Observamos que cuando
la concentracién de NaCl es 0,15 M, que es la concentracién
aproximada del NaCl en el plasma sanguineo (y la de sa-
les de K* en el citoplasma celular) los iones Nat y Cl tienen
por término medio una separacién de 1,9 nm solamente.
Puesto que cada ion Nat y Cl- hidratado posee un diémetro
de 0,5 a 0,7 nm y un agrupamiento tetraédrico de cinco mo-
léculas de agua tiene un didmetro de aproximadamente 0,5 nm,
esta claro que debe haber un cambio considerable en la es-
tructura tridimensional y en las propiedades del agua liquida
cuando se disuelve NaCl en ella a la concentracién aproxi-
mada con que se encuentra en los fluidos biolégicos. Las
sales disueltas tienden, por tanto, a
al atomo de oxigeno de la molécula de agua adyacente a la
cual se halla unido por enlace de hidrégeno, supuesto que
a energia interna de cada molécula sea favorable
HY,
Do-H + On
H
En esta reaccién se producen el ion hidronio (H,O*) y el ion
hidroxilo (OH-). En un litro de agua pura, a 25°C, en un
momento dado existen solamente 1,0 x 10-7 mol de iones H;O*
y una cantidad igual de iones OH’, segin muestran las me-
didas de conductividad eléctrica.
Aunque por convencién, se emplea el simbolo H*, con obje-
to de abreviar, debe recalcarse que en el agua no existen
protones
Amonio
ct
PARTE 1 COMPONENTES MOLECULARES DE LAS CELULAS
racién de un acido, se expresa frente a los equivalentes de
OH> afiadidos, Las formas de tales curvas de valoracién son
muy semejantes para todos los Acidos; la diferencia impor-
tante es que las curvas se hallan desplazadas verticalmente a
Jo largo de la escala de pH. El valor del pH en el punto medio
de la valoracion es numéricamente igual al valor del pK’
del dcido valorado. En el punto medio se hallan presentes
concentraciones equimolares de la especie dadora de protones
(HA) y de Ia aceptora de protones (A-). El pK’ de un acido
puede calcularse, en realidad, a partir del pH en cualquier
punto de la curva de valoracién del acido, siempre que se
conozcan las concentraciones de las especies dadoras y acep-
toras de protones en dicho punto. La forma de la curva de
valoracién puede expresarse por la ecuacién_de Henderson-
Hasselbalch, que es una transformacién logaritmica de la
expresién para la constante de disociacién. Se obtiene como
se indica a continuacion:
(H*)[A7]
THAy
Despejando [H*]:
y tomando el logaritmo negativo de ambos miembros
[HA]
[a]
~ log[H*] =—leg k’ — log
Sustituyendo —log [H*] por pH y —log K’ por pK’
{HA}
HH = pK’ — |
P pl log [A]
Si cambiamos ahora los signos, obtenemos la ecuacién de
Henderson-Hasselbalch:
[A]
[HA]
pH = pK’ + log
cuya forma mas general es la siguiente
, [aceptor de protones]
pH= pK’ + log ‘Tgador de protones]
Esta ecuacién permite el célculo del valor del pK’ de cual-
quier Acido, conociendo la relacién molar de las especies da-
doras y aceptoras de protones, a un pH determinado; permite
también el calculo del pH de un par conjugado acido-basico
que posea un determinado pK’ y una relaci6n molar dada
asimismo permite el calculo de la relacién molar del acido y su
base conjugada si se conocen los correspondientes valores de
pH y pK’. Obsérvese que cuando las concentraciones del dador
y del aceptor de protones son iguales, e! pH observado es nu-
méricamente igual al pK’, La ecuacién de Henderson-Hassel-
balch es fundamental para el tratamiento cuantitativo de todos
Jos equilibrios acido-base en los sistemas biolégicos.
52
Capitulo 2 El agua
Indicadores acido-basicos
El pH de una disolucién puede determinarse mediante el
empleo de colorantes indicadores, la mayor parte de los cuales
son acidos débiles (se designan como HInd). Tales indicado-
res se disocian de acuerdo con el equilibrio
Hind == H* + Ind-
Supongamos que la especie HInd es incolora y que la especie
Ind- es coloreada. La posicién de equilibrio de esta disociacién
y, por tanto, Ia cantidad de luz absorbida por la especie Ind~
a su longitud de onda caracteristica, es determinada por la
concentracién de hidrégeno del medio. En una disolucién
fuertemente acida, el equilibrio se desplaza hacia la izguierda
y se absorbera poca luz a la longitud de onda del maximo de
absorcién de Ind-. En una disolucién basica, la especie Ind-
se halla favorecida, ya que el exceso de iones OH” reacciona
con los iones H* para formar agua, impulsando el equilibrio
hacia Ind-. En este caso se absorbera mucha luz.
‘Tampones
Puede observarse en la figura 2-9 que la curva de valoracién
de cada Acido presenta una zona relativamente llana que se
extiende alrededor de una unidad de pH a cada lado de su
punto medio. En esta zona el pH del sistema cambia relati-
vamente poco cuando se afiaden cantidades mas bien peque-
fias de iones H* 0 OH-. Esta &s la zona en que el par con-
jugado cido-base actiia como fampén; es decir, un sistema
gue tiende a impedir el cambio de pH cuando se afiaden iones
H* o OH-. Para valores de pH fuera de esta zona hay menos
capacidad para resistir los cambios de pH. La capacidad de
tamponamiento es maxima en el mismo punto medio de la
curva de valoracién, es decir, cuando la concentracién de
aceptor de protones se iguala con la de dador de protones,
en la cual se cumple que pH = pK’ (fig. 2-9). La capacidad
de tamponamiento disminuye a medida que aumenta el pH
© desciende, respecto a dicho punto, como consecuencia di-
recta del cambio en la relacién entre las especies aceptora y
dadora de protones. Cada par conjugado acido-basico posee
un pH caracteristico en el cual su capacidad de tampona-
miento es maxima, a saber, el punto en que pH = pK’.
Los fluidos intracelulares y extracelulares de los organis-
mos vivos contienen pares conjugados Acido-basicos, los cuales
actéan como tampones al pH normal de dichos fluidos. El
tampén intracelular mas importante es el par conjugado acido-
basico H,POr-HPO, (pK* = 7,2). Los fosfatos organicos,
tales como la glucosa-6-fosfato y el ATP, contribuyen tam-
bién a la capacidad de tamponamiento en la célula. El prin-
cipal tampén extracelular en la sangre y en los fluidos inters-
ticiales de los vertebrados es el sistema tampén del bicarbo-
nato. La extraordinaria capacidad de tamponamiento del plas-
ma sanguineo puede mostrarse por la siguiente comparacién.
Si se afiade 1 ml de HCI 10 N a 11 de suero fisiol6gico neutro
(aproximadamente 0,15 M en NaCl), el pH de la disolucién
salina descendera hasta pH 2, ya que las disoluciones de
NaCl no poseen capacidad de tamponamiento. Sin embargo,
53
PARTE 1 COMPONENTES MOLECULARES DE LAS CELULAS
si se afiade 1 ml de HCl 10 N a 1 | de plasma sanguineo, el
pH sélo descenderé un poco, desde 7,4 a, aproximadamen-
te, 7,2,
E] sistema tampon del bicarbonato (HyCO;—HCO;) posee
algunos rasgos distintivos. Mientras acttia como tampén, del
mismo modo que otros pares conjugados Acido-basicos, el
pK’ del H,COs, que es un Acido relativamente fuerte, es de
3,8, (tabla 2-7), mucho mas bajo que el intervalo de pH nor~
mal de la sangre. Se plantea la cuestién de cémo un acido
con un valor de pX’ tan bajo es capaz de desempefiar el papel
de tampén fisiolégico a un pH préximo a 7,0. En el sistema
tampén del bicarbonato la especie dadora de protones, el acido
carbénico se halla en equilibrio reversible con el CO; disuelto:
H,CO, == CO,(aq) + H,O
Si tal sistema acuoso esta en contacto con una fase gaseosa,
entonces el anhidrido carbénico disuelto se hallara, a su vez,
en equilibrio entre la fase gaseosa y la acuosa:
CO,(aq) == CO,(g)
Puesto que por la ley de Henry la solubilidad de un gas en’
agua es proporcional a su presin parcial, el pH del sistema
tampén constituido por el bicarbonato es funcién de la presién
parcial del CO; en la fase gaseosa situada sobre la disolucion
tampén. Si aumenta la presién del CO, permaneciendo cons-
tantes las demas variables, el pH del tampén bicarbonato
disminuye, y viceversa. El sistema tampén del bicarbonato
puede regular con eficacia en las proximidades de pH = 7,
en el cual Ia relacién aceptor de protones / dador de protones
es muy elevada, dado que una pequefia cantidad del dador,
el H,CO; se halla en equilibrio inestable con una capacidad
de reserva relativamente grande de gas carbénico en los
pulmones. Bajo cualesquiera condiciones en que la sangre
deba absorber un exceso de iones OH-, el H;CO; empleado
y convertido en HCO; es rapidamente reemplazado por el
CO, gaseoso del gran
S-ar putea să vă placă și
- Cetohexosa de Gran Capacidad EdulcoranteDocument1 paginăCetohexosa de Gran Capacidad EdulcoranteTeresa QuinteroÎncă nu există evaluări
- Propiedades ColigativasDocument51 paginiPropiedades ColigativasJacobo ShasteÎncă nu există evaluări
- Los GlicerolípidosDocument10 paginiLos GlicerolípidosAnonymous x1GAh2pIbÎncă nu există evaluări
- Taller Carbohidratos (Bioquimica)Document18 paginiTaller Carbohidratos (Bioquimica)Dayanna Alexandra Duran Barreiro67% (3)
- Apuntes Biologia Celular y TisularDocument152 paginiApuntes Biologia Celular y TisularFatima Jerez50% (2)
- Cuestionario de LípidosDocument9 paginiCuestionario de LípidosManuelaÎncă nu există evaluări
- Síntesis del colesterol: de la acetil-CoA al mevalonatoDocument2 paginiSíntesis del colesterol: de la acetil-CoA al mevalonatoDvid MedranÎncă nu există evaluări
- Ejercicios Química Orgánica 1Document6 paginiEjercicios Química Orgánica 1Ädriannä_joglärÎncă nu există evaluări
- Ensayo CarbohidratosDocument9 paginiEnsayo CarbohidratosPabloLopez0% (1)
- Glucosa y ácidos orgánicosDocument2 paginiGlucosa y ácidos orgánicosFox gawdÎncă nu există evaluări
- Interacciones Débiles Entre BiomoléculasDocument4 paginiInteracciones Débiles Entre BiomoléculasWolfgang MckenziieÎncă nu există evaluări
- Criofractura técnica microscopíaDocument19 paginiCriofractura técnica microscopíaJuan Jesus Barrios Capuchino0% (1)
- Fórmula QuímicaDocument5 paginiFórmula QuímicaYamileth VillegasÎncă nu există evaluări
- Clase - I - Alcoholes 2Document22 paginiClase - I - Alcoholes 2Yuber DiazÎncă nu există evaluări
- Bioquimica para BachilleratoDocument206 paginiBioquimica para Bachilleratoguianeyav3770100% (5)
- Quimica Del Carbono 2018Document55 paginiQuimica Del Carbono 2018Leandro 05Încă nu există evaluări
- Bioquimica MedicaDocument7 paginiBioquimica Medicayyaraprado785950% (2)
- PROTEÍNASDocument12 paginiPROTEÍNASMarta Sánchez ReinoÎncă nu există evaluări
- Nutrición celular: procesos de obtención de materia y energía por las célulasDocument7 paginiNutrición celular: procesos de obtención de materia y energía por las célulasEdaradna Ybren100% (1)
- Bioquimica GlosarioDocument12 paginiBioquimica GlosarioSalome JuradoÎncă nu există evaluări
- Carotenoides 2011Document31 paginiCarotenoides 2011Ruben Kapa TiconaÎncă nu există evaluări
- Act 14 Quiz FisicoquimicaDocument6 paginiAct 14 Quiz FisicoquimicadacavelÎncă nu există evaluări
- Casos Clinicos - Metabolismo de Lipidos CompletosDocument19 paginiCasos Clinicos - Metabolismo de Lipidos Completosjon diaz100% (1)
- Punto IsoeléctricoDocument7 paginiPunto IsoeléctricoZoiLa BakÎncă nu există evaluări
- LipidosDocument5 paginiLipidosJosé Luis FerrerÎncă nu există evaluări
- Unidad 7 Aldehídos y CetonasDocument26 paginiUnidad 7 Aldehídos y CetonasPablo David PerezÎncă nu există evaluări
- Sintesis de Enalapril PDFDocument4 paginiSintesis de Enalapril PDFRaul Gonzalez100% (1)
- Tipos de Enlaces Interatómicos - 5to SecDocument5 paginiTipos de Enlaces Interatómicos - 5to SecFavio Mauricio GonzaÎncă nu există evaluări
- Libro de La Catedra de BioquimicaDocument211 paginiLibro de La Catedra de BioquimicaCarito GuachoÎncă nu există evaluări
- Qui024 - Ejercicios de Aminoácidos y Péptidos - Semprim2015-3Document12 paginiQui024 - Ejercicios de Aminoácidos y Péptidos - Semprim2015-3Milton Paredes Avalos100% (1)
- Proyecto Educativo Programa de Quimica FarmaceuticaDocument38 paginiProyecto Educativo Programa de Quimica FarmaceuticaeliasgeminisÎncă nu există evaluări
- Metabolismo y Respiración CelularDocument26 paginiMetabolismo y Respiración CelularHerbert JaimesÎncă nu există evaluări
- Guion Reunión Tutor-Padres 2017-8Document17 paginiGuion Reunión Tutor-Padres 2017-8Antonio López FernándezÎncă nu există evaluări
- Actividades Rutas MetabólicasDocument6 paginiActividades Rutas MetabólicasAmador Perez AlejandraÎncă nu există evaluări
- Carbohidratos: estructura, tipos y metabolismoDocument58 paginiCarbohidratos: estructura, tipos y metabolismoLuisianaHEÎncă nu există evaluări
- Bioquímica - EnzimasDocument16 paginiBioquímica - EnzimasCamila Figueroa Abarca100% (1)
- Alcoholes y TiolesDocument56 paginiAlcoholes y TiolesNadia PD100% (1)
- Mol, Osmolaridad, Molalildad y MolaridadDocument5 paginiMol, Osmolaridad, Molalildad y MolaridadSamsaraCT PokémonGo100% (1)
- Examen Bioquimica II MultipleDocument4 paginiExamen Bioquimica II Multiplevalen cristaldoÎncă nu există evaluări
- Lehninger CAPITULO 03Document14 paginiLehninger CAPITULO 03Lyd Granados100% (9)
- OperonDocument2 paginiOperonDaniiel VillegasÎncă nu există evaluări
- Carbohidratos BiologiaDocument23 paginiCarbohidratos BiologiaSearinox21Încă nu există evaluări
- Preguntas y Respuestas UNIDAD II - UNEFMDocument8 paginiPreguntas y Respuestas UNIDAD II - UNEFMIamChachiStephÎncă nu există evaluări
- Cap 12 MetabolismoDocument23 paginiCap 12 MetabolismoPatricia Bustelo100% (1)
- Cadena de Transporte de ElectronesDocument29 paginiCadena de Transporte de ElectronesMarina GrenierÎncă nu există evaluări
- Disacáridos: Sacarosa, Lactosa y MaltosaDocument35 paginiDisacáridos: Sacarosa, Lactosa y MaltosaLuis Alberto Flores ParedesÎncă nu există evaluări
- Problemas MasgoDocument12 paginiProblemas MasgoWenRo Villa AraÎncă nu există evaluări
- Experiencias de GlúcidosDocument47 paginiExperiencias de GlúcidosCarlos Alfredo López0% (1)
- Cadena RespiratoriaDocument2 paginiCadena RespiratoriaJesus RamirezÎncă nu există evaluări
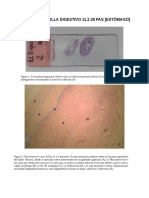
- Informe - Estomago (Tinción PAS)Document3 paginiInforme - Estomago (Tinción PAS)vltg_35643Încă nu există evaluări
- Diseño Molecular de La Vida PDFDocument6 paginiDiseño Molecular de La Vida PDFTania JacintoÎncă nu există evaluări
- Resumen BioquimicaDocument18 paginiResumen BioquimicaMary Elizabeth Bravo Peña100% (2)
- Biología de Smallwood y Green México Primera Edición 1976Document44 paginiBiología de Smallwood y Green México Primera Edición 1976helgvor2608100% (3)
- Classificacio EnzimsDocument3 paginiClassificacio EnzimsjbuedosÎncă nu există evaluări
- El Agua Como BiomoleculaDocument19 paginiEl Agua Como BiomoleculaVictoria TenshiÎncă nu există evaluări
- Función Biológica Del Agua..Document29 paginiFunción Biológica Del Agua..Ricardo Aguado Ornelas0% (2)
- La Estructura Molecular Del AguaDocument13 paginiLa Estructura Molecular Del AguabombmarpÎncă nu există evaluări
- El Agua y La Agua y Estabilidad de Los Alimentos Estabilidad de Los AlimentosDocument28 paginiEl Agua y La Agua y Estabilidad de Los Alimentos Estabilidad de Los AlimentosSarai GarciaÎncă nu există evaluări
- Constantes Fisicas Del Agua y Del HieloDocument17 paginiConstantes Fisicas Del Agua y Del HieloVictor Hugo Carranza Serna100% (1)
- Importancia Biológica Del Agu1Document11 paginiImportancia Biológica Del Agu1Antonio RodriguezÎncă nu există evaluări
- Definicion de Injerto, Colgajo e Implante. Casos Clínicos.Document80 paginiDefinicion de Injerto, Colgajo e Implante. Casos Clínicos.Lyd Granados100% (10)
- Enfermedades Óseas Con Manifestación en Los Maxilares.Document23 paginiEnfermedades Óseas Con Manifestación en Los Maxilares.Lyd Granados100% (4)
- Lehninger CAPITULO 03Document14 paginiLehninger CAPITULO 03Lyd Granados100% (9)
- Manual ColgateDocument76 paginiManual ColgateLyd Granados50% (2)
- Lehninger IntroducciónDocument14 paginiLehninger IntroducciónLyd Granados67% (3)
- Bioquimica de La SalivaDocument2 paginiBioquimica de La SalivaLyd Granados100% (3)
- Farmacologia Clinica para Odontologos Capitulo.Document22 paginiFarmacologia Clinica para Odontologos Capitulo.Lyd Granados75% (8)
- Lehninger CAPITULO 04Document23 paginiLehninger CAPITULO 04Lyd Granados96% (23)
- Apendice DDocument1 paginăApendice DyumesaÎncă nu există evaluări
- Cronología de La BioquímicaDocument6 paginiCronología de La Bioquímicamencho007Încă nu există evaluări
- Injertos Óseos y Su Uso en OdontologíaDocument16 paginiInjertos Óseos y Su Uso en OdontologíaLyd Granados100% (2)
- Clasificación de Películas en EstomatologíaDocument8 paginiClasificación de Películas en EstomatologíaLyd GranadosÎncă nu există evaluări
- Bioquímica Lehninger CAPITULO 01Document21 paginiBioquímica Lehninger CAPITULO 01Lyd Granados75% (8)