S-ar putea să vă placă și
- NEURONUL Reprezinta Unitatea Structural A Si Functional A A Tesutului NervosDocument13 paginiNEURONUL Reprezinta Unitatea Structural A Si Functional A A Tesutului NervoscaltunlilianaÎncă nu există evaluări
- Curs 1 Cergan An 2Document92 paginiCurs 1 Cergan An 2GEORGIANA-CIPRIANA CIUBOTARIUÎncă nu există evaluări
- ADN Replicare Ciclul CelularDocument21 paginiADN Replicare Ciclul CelularCercel Padurariu AlexandraÎncă nu există evaluări
- Sistemul Nervos Vegetativ ExtranevraxialDocument4 paginiSistemul Nervos Vegetativ ExtranevraxialGabriel CiobotaruÎncă nu există evaluări
- Boron Cap 12Document12 paginiBoron Cap 12miaubooksÎncă nu există evaluări
- Diviziunea CelularaDocument44 paginiDiviziunea CelularafbramoÎncă nu există evaluări
- Tesut Conjunctiv Amalinei PDFDocument30 paginiTesut Conjunctiv Amalinei PDFbDashÎncă nu există evaluări
- CelulaDocument15 paginiCelulasebiyÎncă nu există evaluări
- Test Admitere Biologie Cap 18Document7 paginiTest Admitere Biologie Cap 18Elena BordeianuÎncă nu există evaluări
- Celulele ConjunctiveDocument9 paginiCelulele ConjunctiveAriela SăvuţÎncă nu există evaluări
- Completari SNDocument12 paginiCompletari SNCosmina 2510100% (1)
- Sistemul Nervos 3Document4 paginiSistemul Nervos 3Necula100% (2)
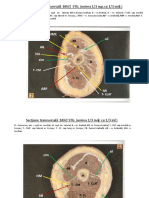
- Sectiuni An.i, Sem.I (1w66754Document10 paginiSectiuni An.i, Sem.I (1w66754Münzer AchtarÎncă nu există evaluări
- Analizatorul KinestezicDocument3 paginiAnalizatorul KinestezicBianca BrlcÎncă nu există evaluări
- Transmit Sensibilități SpecificeDocument4 paginiTransmit Sensibilități SpecificeAlina Simona TeodoroviciÎncă nu există evaluări
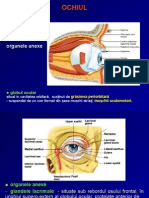
- OchiulDocument31 paginiOchiulRadu Stefan MaricaÎncă nu există evaluări
- Nucleii Din TrunchiDocument3 paginiNucleii Din TrunchiIoana RoşcaÎncă nu există evaluări
- Sistemul Nervos. Generalitati. Neuronul Si SinapsaDocument11 paginiSistemul Nervos. Generalitati. Neuronul Si SinapsaStelian DecebalÎncă nu există evaluări
- Barrons Valori...Document13 paginiBarrons Valori...aida negulescuÎncă nu există evaluări
- Fiziologie TotDocument45 paginiFiziologie TotPasvantu DanielÎncă nu există evaluări
- Benzile Cromozomiale. Tipuri Speciale de CromozomiDocument10 paginiBenzile Cromozomiale. Tipuri Speciale de CromozomiAnda OajdeaÎncă nu există evaluări
- Vascularizatia RinichiuluiDocument2 paginiVascularizatia Rinichiuluilarisa ologuÎncă nu există evaluări
- Test ANALIZATORIDocument11 paginiTest ANALIZATORIRobertina PauletÎncă nu există evaluări
- Tubi UriniferiDocument1 paginăTubi UriniferimarkopavloskiÎncă nu există evaluări
- Inactivarea Cromozomului XDocument7 paginiInactivarea Cromozomului XClaudia SmaduÎncă nu există evaluări
- 4.emisfere CerebraleDocument6 pagini4.emisfere CerebraleMaria MaryÎncă nu există evaluări
- Compuși CarboniliciDocument10 paginiCompuși Carboniliciioana lazaÎncă nu există evaluări
- Cerebelul. Diencefalul. Emisferele CerebraleDocument9 paginiCerebelul. Diencefalul. Emisferele CerebraleVitor MartinsÎncă nu există evaluări
- Pleura Parietala Si VisceralaDocument2 paginiPleura Parietala Si VisceralaFlorin DumitracheÎncă nu există evaluări
- Crosectomia PT VariceDocument7 paginiCrosectomia PT VaricecrisssÎncă nu există evaluări
- Acidul PicricDocument8 paginiAcidul PicricDiana Boşneagă100% (1)
- Analizatorul VizualDocument13 paginiAnalizatorul VizualCosmin MihoreanuÎncă nu există evaluări
- Elementele VitaleDocument12 paginiElementele Vitalevalentina morozovschiÎncă nu există evaluări
- Nucleii Olivari BulbariDocument2 paginiNucleii Olivari BulbariMihai Alexandru ButnariuÎncă nu există evaluări
- Transportul O2 Si CO2Document1 paginăTransportul O2 Si CO2Raluca RadulescuÎncă nu există evaluări
- Modificări Post-TranscriereDocument36 paginiModificări Post-TranscriereAna-Maria IonescuÎncă nu există evaluări
- Lp4-Specializarile Plasmalemei PDFDocument5 paginiLp4-Specializarile Plasmalemei PDFOvidiu MarianÎncă nu există evaluări
- Ventriculi LateraliDocument4 paginiVentriculi LateraliBiancaÎncă nu există evaluări
- Histologie - Os Partea IIDocument7 paginiHistologie - Os Partea IIMiruna AndreeaÎncă nu există evaluări
- Notițe Admitere Medicină: 2020 Vol. IIIDocument108 paginiNotițe Admitere Medicină: 2020 Vol. IIIAndrada UngureanÎncă nu există evaluări
- Fiziologia Respiratiei IIIDocument39 paginiFiziologia Respiratiei IIIMădă Iorga100% (1)
- Anatomie CURS 7Document4 paginiAnatomie CURS 7Elena ElenaÎncă nu există evaluări
- CEREBELULDocument6 paginiCEREBELULSabina SabinaaÎncă nu există evaluări
- Tiamina: 1 Structura ChimicaDocument10 paginiTiamina: 1 Structura ChimicaDaniela DarabanÎncă nu există evaluări
- 3., - ŢesuturileDocument5 pagini3., - ŢesuturileDiana Petrea100% (1)
- Structura Si Functiile Neuronului - SinapseDocument3 paginiStructura Si Functiile Neuronului - SinapseMihaela CenușeÎncă nu există evaluări
- CURS 12 Compusi DicarboniliciDocument10 paginiCURS 12 Compusi DicarboniliciMerla OanaÎncă nu există evaluări
- Tehnici de Colorare in Biologia CelularaDocument11 paginiTehnici de Colorare in Biologia CelularaAlexandra ManuelaÎncă nu există evaluări
- Subiecte Rezolvate ColocviuDocument18 paginiSubiecte Rezolvate ColocviuCristina IoanaÎncă nu există evaluări
- Fiziologia Muschiului - 2019Document43 paginiFiziologia Muschiului - 2019Narcis MessiÎncă nu există evaluări
- 2008 Biologie Etapa Judeteana Subiecte Clasa A XI-A 0Document11 pagini2008 Biologie Etapa Judeteana Subiecte Clasa A XI-A 0classicgrin100% (1)
- LP 2 Analiza Cromozomilor Sexuali La OmDocument3 paginiLP 2 Analiza Cromozomilor Sexuali La OmAna Maria DanÎncă nu există evaluări
- Analizatorul Olfactiv Si GustativDocument4 paginiAnalizatorul Olfactiv Si GustativDiana CismaruÎncă nu există evaluări
- FiziopatologieDocument30 paginiFiziopatologieanne9oÎncă nu există evaluări
- Subiecte Rezolvate SNCDocument5 paginiSubiecte Rezolvate SNCsirbumÎncă nu există evaluări
- Tesut NervosDocument11 paginiTesut NervosSimion CristianÎncă nu există evaluări
- Structura Morfologică A NeuronuluiDocument4 paginiStructura Morfologică A NeuronuluiDiana Florentina AdumitrăceseiÎncă nu există evaluări
- HISTO CURS 13 - Review PDFDocument13 paginiHISTO CURS 13 - Review PDFSerban ElenaÎncă nu există evaluări
- Sistemul Nervos SomaticDocument16 paginiSistemul Nervos SomaticAna Karisia67% (3)
- L 3 - Sistemul Nervos (Introducere, Maduva Spinarii)Document10 paginiL 3 - Sistemul Nervos (Introducere, Maduva Spinarii)Svetlana Doibani100% (1)
- Concl UziDocument1 paginăConcl UziAlamentheÎncă nu există evaluări
- CursDocument42 paginiCursAlamentheÎncă nu există evaluări
- Simpaticul LombarDocument2 paginiSimpaticul LombarAlamentheÎncă nu există evaluări
- Bacterio Pe Sub 1Document41 paginiBacterio Pe Sub 1AlamentheÎncă nu există evaluări
- Peretii AbdomenuluiDocument13 paginiPeretii AbdomenuluiFlorin TeodorescuÎncă nu există evaluări
- Sectiunea 24 Faringe LaringeDocument8 paginiSectiunea 24 Faringe LaringeblyblyÎncă nu există evaluări
- Subiecte Anato Sem I An 2Document3 paginiSubiecte Anato Sem I An 2AlamentheÎncă nu există evaluări
- Culegere de Teste Grila Iasi 2013Document151 paginiCulegere de Teste Grila Iasi 2013Adina Cristina FriciuÎncă nu există evaluări