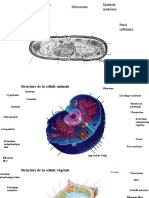
S-ar putea să vă placă și
- 6-Reticulum Endoplasmique-Corps de GolgiSANSannexe2019Document41 pagini6-Reticulum Endoplasmique-Corps de GolgiSANSannexe2019samÎncă nu există evaluări
- Applications de la spectrophotomérie en phytochimie: sciencesDe la EverandApplications de la spectrophotomérie en phytochimie: sciencesÎncă nu există evaluări
- 9 CLASSE DE Tle C-1Document103 pagini9 CLASSE DE Tle C-1Njiasse MaazouÎncă nu există evaluări
- Le Systeme Endo MembranaireDocument30 paginiLe Systeme Endo MembranaireAbd El AzizÎncă nu există evaluări
- Bio CellulaireDocument44 paginiBio CellulaireTouri Na100% (1)
- Le Tissu EpithelialDocument8 paginiLe Tissu EpithelialFaculté De Médecine BécharÎncă nu există evaluări
- Evaluation - Examens Finaux - Examen Final 2000 (Corrigé)Document11 paginiEvaluation - Examens Finaux - Examen Final 2000 (Corrigé)mounaÎncă nu există evaluări
- Unité 1 - La Cellule, Unité Du VivantDocument8 paginiUnité 1 - La Cellule, Unité Du VivantIrene PGÎncă nu există evaluări
- Ctenaires PDFDocument13 paginiCtenaires PDFNsangwaÎncă nu există evaluări
- Chap1 EucaryoteDocument4 paginiChap1 Eucaryoteslimdamak100% (1)
- TP N°3 Les ParenchymesDocument10 paginiTP N°3 Les ParenchymesSofiane BousifeÎncă nu există evaluări
- TD CorrigeDocument5 paginiTD Corrigeam47Încă nu există evaluări
- Embryologie s1Document127 paginiEmbryologie s1MeRyem El MejdoubÎncă nu există evaluări
- Cyto1an-Systeme EndomembranaireDocument82 paginiCyto1an-Systeme EndomembranaireAdam FellousÎncă nu există evaluări
- Cours Annuel SVT EDUCCIADocument96 paginiCours Annuel SVT EDUCCIAYvanna TebeuÎncă nu există evaluări
- Chapitre III La FécondationDocument7 paginiChapitre III La FécondationDel HadÎncă nu există evaluări
- 2021 - Cours La Cellule Version ComplèteDocument10 pagini2021 - Cours La Cellule Version Complèteagathe surusÎncă nu există evaluări
- Résumé ÉpithéliumDocument6 paginiRésumé ÉpithéliumAmiddoÎncă nu există evaluări
- TD HistoDocument2 paginiTD HistoAbdul Koudouss Kone100% (1)
- Développement EmbryonnaireDocument9 paginiDéveloppement EmbryonnaireEva Peignier100% (1)
- TD Bio Cell 1ère MedDocument65 paginiTD Bio Cell 1ère Medouma imaÎncă nu există evaluări
- Histo1an-Epitheliums RevetementDocument64 paginiHisto1an-Epitheliums RevetementZÏzoo MamÏzooÎncă nu există evaluări
- B.A Les VertebresDocument61 paginiB.A Les Vertebresimane chakibÎncă nu există evaluări
- Intro Sur Histo VegetaleDocument6 paginiIntro Sur Histo Vegetaleشكوب ستانÎncă nu există evaluări
- Membrane PlasmiqueDocument53 paginiMembrane Plasmiqueyamina kadi100% (1)
- Cours de Tle C & TI - Séquence 1Document5 paginiCours de Tle C & TI - Séquence 1Romario nzou100% (1)
- La Cellule EucaryoteDocument9 paginiLa Cellule Eucaryoteherrere64Încă nu există evaluări
- Tle D SVTEEHBDocument6 paginiTle D SVTEEHBJOEL LOIC100% (1)
- Fiches Enseignement ScientifiqueDocument15 paginiFiches Enseignement ScientifiquenoorÎncă nu există evaluări
- Bryophytes 1Document6 paginiBryophytes 1sumaleeÎncă nu există evaluări
- 7 Les Epitheliums de RevetementDocument6 pagini7 Les Epitheliums de Revetementlyna7otsÎncă nu există evaluări
- TP1 de CytologieDocument6 paginiTP1 de CytologieSonja BendadaÎncă nu există evaluări
- Cours Biologie Végétale Les CormophytesDocument194 paginiCours Biologie Végétale Les Cormophyteskhadijasaadlah74Încă nu există evaluări
- TD2 Exercice 1:: Module: Physiologie de DigestionDocument2 paginiTD2 Exercice 1:: Module: Physiologie de DigestionImi BenÎncă nu există evaluări
- 1ere S1 Physiologie - LE MILIEU INTERIEUR DR Rouatbi SoniaDocument125 pagini1ere S1 Physiologie - LE MILIEU INTERIEUR DR Rouatbi Sonianourhene rakroukiÎncă nu există evaluări
- Devoir de Biologie CellulaireDocument1 paginăDevoir de Biologie CellulaireMounir Benamara100% (1)
- Partie I Chap I Le Milieu InterieurDocument15 paginiPartie I Chap I Le Milieu Interieurslimdamak0% (1)
- La CelluleDocument52 paginiLa Cellulesimo.elhaouate.1000Încă nu există evaluări
- Cour SVT TleDDocument122 paginiCour SVT TleDNdjidama youssoufa0% (1)
- TRAVAUX DIRIGE DE BOA 356 (DR Wankeu)Document11 paginiTRAVAUX DIRIGE DE BOA 356 (DR Wankeu)Staline DonkengÎncă nu există evaluări
- Cours Milieu Intérieur 2020Document34 paginiCours Milieu Intérieur 2020Dan MayaÎncă nu există evaluări
- Gamatogenese Et FecondationDocument47 paginiGamatogenese Et FecondationAmïn Fïlalï100% (1)
- Tweet: La Synthèse Des ProtéinesDocument59 paginiTweet: La Synthèse Des ProtéinesNadaÎncă nu există evaluări
- TP Tissu MusculaireDocument5 paginiTP Tissu Musculaireyacine bounamaÎncă nu există evaluări
- Compartiment LiquidienDocument9 paginiCompartiment LiquidienFofana LancinéÎncă nu există evaluări
- CytosqueletteDocument24 paginiCytosquelettefellah aminaÎncă nu există evaluări
- Présentation.41.ppt PAROIDocument12 paginiPrésentation.41.ppt PAROIRêda SlimiÎncă nu există evaluări
- ARTHROPODESDocument8 paginiARTHROPODESThug FamilyÎncă nu există evaluări
- Cours No 02-La CelluleDocument13 paginiCours No 02-La CelluleFeriel FerielÎncă nu există evaluări
- La Cellule Bactérienne: Morphologie Et Structure: 1. TaxonomieDocument9 paginiLa Cellule Bactérienne: Morphologie Et Structure: 1. Taxonomiekyara turpinÎncă nu există evaluări
- BIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFDocument81 paginiBIO 224 Anatomie - Comparée (2020) - Compressé - 0 PDFTONDI ABDOU CHACOUR67% (3)
- Les Acides NucléiquesDocument22 paginiLes Acides NucléiquesMahmoudane TouréÎncă nu există evaluări
- Cours BV - MSVT2 - PDF Cryptogamie PDFDocument131 paginiCours BV - MSVT2 - PDF Cryptogamie PDFElbouk Valia100% (2)
- Milieu Intérieur de L'organismeDocument88 paginiMilieu Intérieur de L'organismeMoussaba DoumbouyaÎncă nu există evaluări
- La Chaine AlimentaireDocument14 paginiLa Chaine AlimentaireAhmed BoussabehÎncă nu există evaluări
- 1 - Organisation Générale de La CelluleDocument6 pagini1 - Organisation Générale de La Cellulebaya100% (1)
- Le Cytosquelette. 2021 CH7Document68 paginiLe Cytosquelette. 2021 CH7paul destinÎncă nu există evaluări
- Histo EpitheliumDocument13 paginiHisto EpitheliumMeriem Ab100% (3)
- QUESTIONNAIRE COURS BIOLOGIE CELLULAIRE 1ERE ANNEE PHARMACIE - No KeyDocument11 paginiQUESTIONNAIRE COURS BIOLOGIE CELLULAIRE 1ERE ANNEE PHARMACIE - No KeyKeishaÎncă nu există evaluări
- BTS Dietetique Cellules Eucaryotes Et ProcaryotesDocument2 paginiBTS Dietetique Cellules Eucaryotes Et ProcaryotesEkissi Etienne OrsotÎncă nu există evaluări
- Bilogie CoursDocument6 paginiBilogie CoursAziz Chaieb100% (1)
- Cours BIO-1-Définition & Origine de La VieDocument90 paginiCours BIO-1-Définition & Origine de La VieBenjamin Plee100% (1)
- Khanfri 1-200Document22 paginiKhanfri 1-200Neda ArÎncă nu există evaluări
- Chapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8Document13 paginiChapitre VII Le Reticulum Endoplasmique Et Lappareil de Golgi B8kim namjoonÎncă nu există evaluări
- Travaux Dirigés Biologie Cellulaire Corrigé 2023Document47 paginiTravaux Dirigés Biologie Cellulaire Corrigé 2023Adamou Aboubé AbdoulayeÎncă nu există evaluări
- A LégenderDocument3 paginiA LégenderFatima Ezzahra AchkouÎncă nu există evaluări
- Ultrastructure Composition Chimique Et Roles de Constituants CellulaireDocument4 paginiUltrastructure Composition Chimique Et Roles de Constituants Cellulairefyvsgbj92bÎncă nu există evaluări
- PosterDocument5 paginiPosterBrahim OuarhlentÎncă nu există evaluări
- Membrane PlasmiqueDocument31 paginiMembrane PlasmiqueMichael GuillienÎncă nu există evaluări
- C Proc Et EucDocument23 paginiC Proc Et EucAya youyaÎncă nu există evaluări
- Partie 2. Cours de BioCell 1ASNVDocument17 paginiPartie 2. Cours de BioCell 1ASNVايمن معطااللهÎncă nu există evaluări
- Annoter Le Schéma en Cliquant Sur Le Mot Ou LDocument4 paginiAnnoter Le Schéma en Cliquant Sur Le Mot Ou LKéléfaÎncă nu există evaluări
- Cours BioCell Chap1et2Document107 paginiCours BioCell Chap1et2NancyÎncă nu există evaluări
- Les Cellules Eucaryotes 2020 - 2021Document5 paginiLes Cellules Eucaryotes 2020 - 2021Jimingot jams0% (1)
- La BiosphèreDocument19 paginiLa Biosphèredam ayusoÎncă nu există evaluări
- Biologie CellulaireDocument26 paginiBiologie CellulaireBourobou bourobou Chadrack gaelÎncă nu există evaluări
- Ladressage Des Protéines - PDF Dans RERDocument48 paginiLadressage Des Protéines - PDF Dans RERamal ELYOUBIÎncă nu există evaluări
- Biologie Cellulaire - PR KHANFRI Jamal Eddine - 16102020 - Cours Texte PDFDocument118 paginiBiologie Cellulaire - PR KHANFRI Jamal Eddine - 16102020 - Cours Texte PDFRania MaddahÎncă nu există evaluări
- La CytologieDocument97 paginiLa CytologieelohimmvulaÎncă nu există evaluări
- Cours Systeme EndomembranaireDocument9 paginiCours Systeme EndomembranaireIOana Borca100% (1)
- Cyto1an-Systeme EndomembranaireDocument82 paginiCyto1an-Systeme EndomembranaireAdam FellousÎncă nu există evaluări
- COURS BC - Système EndomembranaireDocument30 paginiCOURS BC - Système EndomembranaireMohammed El MachmoulÎncă nu există evaluări
- Partie 1. Cours de BioCell 1ASNVDocument9 paginiPartie 1. Cours de BioCell 1ASNVايمن معطااللهÎncă nu există evaluări
- Cours de Biologie Cellulaire Rabat 2017-2018Document98 paginiCours de Biologie Cellulaire Rabat 2017-2018Badr ZbidaÎncă nu există evaluări
- 3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Document56 pagini3.schema General Des Cellules Procaryotes Et Eucaryotes 2021-2022Le BoyaïsteÎncă nu există evaluări
- 101à200 QCM FMA1 Bio CellDocument12 pagini101à200 QCM FMA1 Bio CellFéz EyÎncă nu există evaluări
- Programmes PASS 2021 2022 Majeure SanteDocument11 paginiProgrammes PASS 2021 2022 Majeure SanteOmas MontpellierÎncă nu există evaluări