S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- CFA L1 Ethics Questions and AnswersDocument94 paginiCFA L1 Ethics Questions and AnswersMaulik PatelÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Caracterisation D'une Entreprise anglosaxonne-ETLV - STMG1.Document2 paginiCaracterisation D'une Entreprise anglosaxonne-ETLV - STMG1.meredith.licagaÎncă nu există evaluări
- Sample Database of SQL in Mysql FormatDocument7 paginiSample Database of SQL in Mysql FormatsakonokeÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- ECDIS Presentation Library 4Document16 paginiECDIS Presentation Library 4Orlando QuevedoÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- MKT STGDocument106 paginiMKT STGAmeya Singh RajputÎncă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Rated Operational Current: InstructionsDocument12 paginiRated Operational Current: InstructionsJhon SanabriaÎncă nu există evaluări
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- VISCOLAM202 D20 Acrylic 20 Thickeners 202017Document33 paginiVISCOLAM202 D20 Acrylic 20 Thickeners 202017Oswaldo Manuel Ramirez MarinÎncă nu există evaluări
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Lab 08: SR Flip Flop FundamentalsDocument6 paginiLab 08: SR Flip Flop Fundamentalsjitu123456789Încă nu există evaluări
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Inkolo Namasiko Kuyamakha Umuntu - Brainly - inDocument1 paginăInkolo Namasiko Kuyamakha Umuntu - Brainly - inxqxfkqpy5qÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Community Reinvestment Act in The Age of Fintech and Bank CompetitionDocument28 paginiThe Community Reinvestment Act in The Age of Fintech and Bank CompetitionHyder AliÎncă nu există evaluări
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- 20131022-Additive Manufacturing & Allied Technologies, PuneDocument56 pagini20131022-Additive Manufacturing & Allied Technologies, Puneprakush_prakushÎncă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Consumer Price SummaryDocument5 paginiConsumer Price SummaryKJ HiramotoÎncă nu există evaluări
- Eat Something DifferentDocument3 paginiEat Something Differentsrajendr200100% (1)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Genstat Release 10.3de (Pc/Windows 7) 28 May 2012 06:35:59Document6 paginiGenstat Release 10.3de (Pc/Windows 7) 28 May 2012 06:35:59Anna Nur HidayatiÎncă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Cover Sheet: Online Learning and Teaching (OLT) Conference 2006, Pages Pp. 21-30Document12 paginiCover Sheet: Online Learning and Teaching (OLT) Conference 2006, Pages Pp. 21-30Shri Avinash NarendhranÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Read The Text and Answer The QuestionsDocument5 paginiRead The Text and Answer The QuestionsDanny RuedaÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- Corporate Governance Guidelines GMDocument15 paginiCorporate Governance Guidelines GMWaqas MahmoodÎncă nu există evaluări
- CHAPTER I KyleDocument13 paginiCHAPTER I KyleCresiel Pontijon100% (1)
- Ansi Numerical CodeDocument6 paginiAnsi Numerical Codekachra13Încă nu există evaluări
- 1and5.microscopes, Specializedstem Cells, Homeostasis - Answer KeyDocument1 pagină1and5.microscopes, Specializedstem Cells, Homeostasis - Answer KeyMCarmen López CastroÎncă nu există evaluări
- Tutorial 4 QuestionsDocument3 paginiTutorial 4 QuestionshrfjbjrfrfÎncă nu există evaluări
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
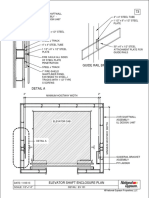
- Guide Rail Bracket AssemblyDocument1 paginăGuide Rail Bracket AssemblyPrasanth VarrierÎncă nu există evaluări
- Week 1 Lab #2 - Microscopy & Microscopic Examination of Living MicroorganismsDocument53 paginiWeek 1 Lab #2 - Microscopy & Microscopic Examination of Living MicroorganismsNgoc PhamÎncă nu există evaluări
- Dr. Muhammad Mumtaz: Thesis TitleDocument10 paginiDr. Muhammad Mumtaz: Thesis TitleDr. Muhammad MumtazÎncă nu există evaluări
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- RISO RZ User GuideDocument112 paginiRISO RZ User GuideJojo AritallaÎncă nu există evaluări
- Veris Case StudyDocument2 paginiVeris Case StudyPankaj GargÎncă nu există evaluări
- Karamadi - A Waning Practice of Shore Seine Operation Along Kerala, SouthwestDocument6 paginiKaramadi - A Waning Practice of Shore Seine Operation Along Kerala, SouthwestQ8123Încă nu există evaluări
- Annexure I Project Details DateDocument4 paginiAnnexure I Project Details DateAshish SinghaniaÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (120)
- Oracle® Secure Backup: Installation and Configuration Guide Release 10.4Document178 paginiOracle® Secure Backup: Installation and Configuration Guide Release 10.4andrelmacedoÎncă nu există evaluări
- Fuel Injection PDFDocument11 paginiFuel Injection PDFscaniaÎncă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)