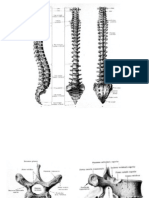
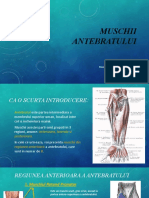
S-ar putea să vă placă și
- MuscularDocument9 paginiMuscularpufdepapadieÎncă nu există evaluări
- Referat FiziollogieDocument7 paginiReferat FiziollogiedarlaloredanaÎncă nu există evaluări
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Paralizia de Plex BrahialDocument16 paginiParalizia de Plex BrahialRucsandra AvirvareiÎncă nu există evaluări
- Motilitate ReflexeDocument9 paginiMotilitate ReflexeNorbert StenczelÎncă nu există evaluări
- Traumatismele Maduvei SpinariiDocument27 paginiTraumatismele Maduvei SpinariiIonut Si Madalina PatrascuÎncă nu există evaluări
- Leziunile de Nervi Periferici Ghid de Diagnostic Si TratamentDocument85 paginiLeziunile de Nervi Periferici Ghid de Diagnostic Si Tratamentizo_katenaÎncă nu există evaluări
- C 5 - Masaj Clasic Fasc 5Document8 paginiC 5 - Masaj Clasic Fasc 5Vornicu OanaÎncă nu există evaluări
- Sindromul de Neuron Motor Central (Sindromul Piramidal) Curs4Document7 paginiSindromul de Neuron Motor Central (Sindromul Piramidal) Curs4Cristina-Marinela ObrejaÎncă nu există evaluări
- Atrofia Musculară SpinalăDocument10 paginiAtrofia Musculară SpinalăacbernevigÎncă nu există evaluări
- 34.tumorile Osului - DR - Gonta AndreiDocument17 pagini34.tumorile Osului - DR - Gonta AndreiBeatrice CioroiuÎncă nu există evaluări
- Leziunile Nervilor PerifericiDocument8 paginiLeziunile Nervilor PerifericiLauraSÎncă nu există evaluări
- ObosealaDocument35 paginiObosealaClaudia Nita100% (1)
- Patologia Maduvei SpinariiDocument36 paginiPatologia Maduvei SpinariiIoanaÎncă nu există evaluări
- Examen Neuro PDFDocument87 paginiExamen Neuro PDFCarolinaZloiÎncă nu există evaluări
- Coada de CalDocument4 paginiCoada de CalAdi RughiÎncă nu există evaluări
- Tesutul MuscularDocument10 paginiTesutul MuscularGiku MikushimirikuÎncă nu există evaluări
- Ashworth PennDocument1 paginăAshworth Penncelena8Încă nu există evaluări
- Tehnici de Facilitare Neuromusculară ProprioceptivăDocument8 paginiTehnici de Facilitare Neuromusculară ProprioceptivăB.Sz.Încă nu există evaluări
- Sindromul de Neuron Motor de Tip Central Si PerifericDocument5 paginiSindromul de Neuron Motor de Tip Central Si PerifericAlexandra G. Cazan100% (1)
- Curs Neuro 2 2010Document40 paginiCurs Neuro 2 2010Ciprian Cozmuleasa100% (3)
- Caracteristici Morfofunctionale Ale Aparatului LocomotorDocument4 paginiCaracteristici Morfofunctionale Ale Aparatului LocomotorSilviu LipanÎncă nu există evaluări
- Leziuni Musculare de SuprasolicitareDocument16 paginiLeziuni Musculare de SuprasolicitareValentin Bogdan VfbÎncă nu există evaluări
- Neurologie Si Neurochirurgie Anul 4Document3 paginiNeurologie Si Neurochirurgie Anul 4Nadea TomshaÎncă nu există evaluări
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- Tabele EvaluareDocument12 paginiTabele EvaluareDuta DanielaÎncă nu există evaluări
- Itr BFKT LP 04-13Document46 paginiItr BFKT LP 04-13Alin IonutÎncă nu există evaluări
- Sindromul de Neuron Motor CentralDocument9 paginiSindromul de Neuron Motor CentralHagima Adrian-MihaiÎncă nu există evaluări
- HidrocefaliaDocument37 paginiHidrocefaliacristiana100% (1)
- Principiile Morale Ale KinetoterapeutuluiDocument16 paginiPrincipiile Morale Ale KinetoterapeutuluiDaniela Paduraru100% (1)
- Gabat 1Document165 paginiGabat 1andrei_costachiÎncă nu există evaluări
- RELAXAREADocument34 paginiRELAXAREAAdrian VladÎncă nu există evaluări
- Sistem MuschiDocument19 paginiSistem MuschiDan MassonÎncă nu există evaluări
- C8 AlgoneurodistrofiaDocument8 paginiC8 Algoneurodistrofiatzimboaka .tzimbyÎncă nu există evaluări
- Gametogeneza. Ovulatie. Fecundatie. Nidatie. Placentatia. Fiziologia PlacenteiDocument15 paginiGametogeneza. Ovulatie. Fecundatie. Nidatie. Placentatia. Fiziologia PlacenteiIna SimacheÎncă nu există evaluări
- Prezentare CerebelosDocument28 paginiPrezentare CerebelosTeona OanaÎncă nu există evaluări
- Osteologie Si ArtrologieDocument51 paginiOsteologie Si ArtrologieCristina IoanaÎncă nu există evaluări
- Evaluarea Tolerantei La EfortDocument11 paginiEvaluarea Tolerantei La EfortRoxana BuculescuÎncă nu există evaluări
- 5 HtaDocument41 pagini5 HtaGhica CostinÎncă nu există evaluări
- Plexurile NervoaseDocument4 paginiPlexurile NervoaseCristian GrecuÎncă nu există evaluări
- Coloana CervicalaDocument27 paginiColoana CervicalaValentin Bogdan VfbÎncă nu există evaluări
- 3 Sindromul Cerebelos Si Sindromul VestibularDocument7 pagini3 Sindromul Cerebelos Si Sindromul VestibulardcnÎncă nu există evaluări
- Metodologia Antrenamentului Personalizat Curs PDFDocument58 paginiMetodologia Antrenamentului Personalizat Curs PDFmihaelaboncataÎncă nu există evaluări
- Semiologie NeurologicaDocument14 paginiSemiologie NeurologicaAlin OprisanÎncă nu există evaluări
- Biomecanica Intrebari Si RaspunsuriDocument34 paginiBiomecanica Intrebari Si RaspunsuriAlexandra MarinÎncă nu există evaluări
- Biomecanica Piciorului Dr. Traian UrsuDocument14 paginiBiomecanica Piciorului Dr. Traian UrsuTraian UrsuÎncă nu există evaluări
- Fiziologie SNCDocument63 paginiFiziologie SNCMihaiCristianÎncă nu există evaluări
- Sindromul RadicularDocument14 paginiSindromul RadicularIonelLucianStanÎncă nu există evaluări
- Muschii Antebratului Partea AnterioaraDocument10 paginiMuschii Antebratului Partea AnterioaraDelia-Rafaela CucÎncă nu există evaluări
- Aplicatiile Masajului in SportDocument7 paginiAplicatiile Masajului in SportDraghici ValentinÎncă nu există evaluări
- SDR - PiramidalDocument44 paginiSDR - PiramidalHussein TfÎncă nu există evaluări
- Examen Neurologic 33Document1 paginăExamen Neurologic 33Alexandra MardareÎncă nu există evaluări
- Curs Neuromuscular-Radu BragaDocument14 paginiCurs Neuromuscular-Radu BragaVicențiu Răzvan VornicuÎncă nu există evaluări
- Fiziologia Contractiei MusculareDocument4 paginiFiziologia Contractiei MusculareIoana irimiaÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- Curs 4 FiziologieDocument10 paginiCurs 4 FiziologieDAROIU REBECAGEORGIANAÎncă nu există evaluări
- Biofizica Aparatului LocomotorDocument10 paginiBiofizica Aparatului LocomotorAurel BoleaÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- HIPOTIROIDISMDocument22 paginiHIPOTIROIDISMIosca-Antonesei Oana100% (4)
- Lori SepsisDocument5 paginiLori SepsisLoredana BoghezÎncă nu există evaluări
- Formular de Renuntare Din ContractDocument1 paginăFormular de Renuntare Din ContractLoredana BoghezÎncă nu există evaluări
- Tulbur Hidrice Si ElectroliticeDocument12 paginiTulbur Hidrice Si ElectroliticeMaria Amelia MercaÎncă nu există evaluări
- FIZIOPATOLOGIEDocument23 paginiFIZIOPATOLOGIELoredana BoghezÎncă nu există evaluări
- Anexa 2. Chestionar - Studenti Grad de SatisfactieDocument3 paginiAnexa 2. Chestionar - Studenti Grad de SatisfactieBanariu Gheorghe MihailÎncă nu există evaluări
- Scleroza MultiplăDocument10 paginiScleroza MultiplăLoredana BoghezÎncă nu există evaluări
- Biostatistica 2Document16 paginiBiostatistica 2Loredana BoghezÎncă nu există evaluări
- Artroplastie FINAL PT EvaluatoriDocument9 paginiArtroplastie FINAL PT EvaluatoriLoredana BoghezÎncă nu există evaluări
- BayeDocument71 paginiBayeLoredana BoghezÎncă nu există evaluări
- PediatrieDocument100 paginiPediatrieFlorin Ardeleanu100% (1)
- 8 - Cateterismul Cardiac - Dr. Nicolae FlorescuDocument10 pagini8 - Cateterismul Cardiac - Dr. Nicolae Florescublu_bee100% (1)
- Scrisoare de Intenție Maria PușcașuDocument3 paginiScrisoare de Intenție Maria PușcașuLoredana BoghezÎncă nu există evaluări
- Licenta Cardio Lore SecundDocument84 paginiLicenta Cardio Lore SecundLoredana BoghezÎncă nu există evaluări
- Evaluarea Ecocardiografică În Insuficiența Cardiacă Acută Și În Edemul Pulmonar AcutDocument12 paginiEvaluarea Ecocardiografică În Insuficiența Cardiacă Acută Și În Edemul Pulmonar AcutLoredana BoghezÎncă nu există evaluări
- Sufletul Neamului Romanesc Vol 2 - Dacia Felix, Augusta, Terra Mater Sau Pamantul FagaduinteiDocument121 paginiSufletul Neamului Romanesc Vol 2 - Dacia Felix, Augusta, Terra Mater Sau Pamantul Fagaduinteiraluca17oct100% (1)
- PediatrieDocument100 paginiPediatrieFlorin Ardeleanu100% (1)
- Refractia OcularaDocument8 paginiRefractia OcularaanymaryaÎncă nu există evaluări
- Cuprins LicentaDocument4 paginiCuprins LicentaLoredana BoghezÎncă nu există evaluări
- Licenta Iulia Iordache PDFDocument86 paginiLicenta Iulia Iordache PDFLoredana BoghezÎncă nu există evaluări
- Variabile Teste VHMBM NMNDocument8 paginiVariabile Teste VHMBM NMNMădălina MateiÎncă nu există evaluări
- Plan Managerial Maria PușcașuDocument4 paginiPlan Managerial Maria PușcașuLoredana BoghezÎncă nu există evaluări
- Scleroza MultiplăDocument10 paginiScleroza MultiplăLoredana BoghezÎncă nu există evaluări
- Tulbur Hidrice Si ElectroliticeDocument12 paginiTulbur Hidrice Si ElectroliticeMaria Amelia MercaÎncă nu există evaluări
- Scrisoare de Intenție Maria PușcașuDocument3 paginiScrisoare de Intenție Maria PușcașuLoredana BoghezÎncă nu există evaluări
- Baza Date FiaDocument33 paginiBaza Date FiaLoredana BoghezÎncă nu există evaluări
- 08 RaaDocument46 pagini08 RaaGrigore AlexandraÎncă nu există evaluări
- Scrisoare de Intenție Maria PușcașuDocument3 paginiScrisoare de Intenție Maria PușcașuLoredana BoghezÎncă nu există evaluări
- 08 RaaDocument46 pagini08 RaaGrigore AlexandraÎncă nu există evaluări
- 09 Cardiopatii CongenitaleDocument60 pagini09 Cardiopatii CongenitaleLoredana BoghezÎncă nu există evaluări