S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (72)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Qubit 4 Fluorometer-Fast Answers For Precious Samples: Find Out If You Have Enough DNA or RNA For Your ExperimentDocument2 paginiQubit 4 Fluorometer-Fast Answers For Precious Samples: Find Out If You Have Enough DNA or RNA For Your ExperimentmisuakechiÎncă nu există evaluări
- Mccee NotesDocument3 paginiMccee Notesblueivy66Încă nu există evaluări
- Mechanisms of Bacterial PathogenesisDocument34 paginiMechanisms of Bacterial PathogenesisMamta ShindeÎncă nu există evaluări
- Hiv-1 Quant PCRDocument62 paginiHiv-1 Quant PCRyousrazeidan1979Încă nu există evaluări
- Fish TishDocument151 paginiFish Tishabhinav_choudhary03Încă nu există evaluări
- Mengeles ChildrenDocument3 paginiMengeles Childrenapi-309154321Încă nu există evaluări
- HomeostasisDocument13 paginiHomeostasisAkeem K JacksonÎncă nu există evaluări
- Building On Success: A Bright Future For Peptide TherapeuticsDocument7 paginiBuilding On Success: A Bright Future For Peptide TherapeuticsDiana PachónÎncă nu există evaluări
- Biotechnology: Quarter 3 - Module 2: Genetic ManipulationDocument24 paginiBiotechnology: Quarter 3 - Module 2: Genetic ManipulationJohn Mark Laurio82% (17)
- Genomics of Plants and Fungi - Prade and BohnertDocument395 paginiGenomics of Plants and Fungi - Prade and BohnertFernanda CastroÎncă nu există evaluări
- The Reproductive SystemDocument119 paginiThe Reproductive SystemAshis karmakar100% (10)
- The Benefits of Alpha Lipoic Acid: A Potent AntioxidantDocument56 paginiThe Benefits of Alpha Lipoic Acid: A Potent AntioxidantAisya Amalia MuslimaÎncă nu există evaluări
- Stomatognathic SystemDocument129 paginiStomatognathic Systemsurya kingsly100% (1)
- ACTN3 Genotype Is Associated With Human Elite Athletic PerformanceDocument5 paginiACTN3 Genotype Is Associated With Human Elite Athletic PerformanceLuciano JuniorÎncă nu există evaluări
- Differential Diagnosis and Comorbidity Distinguishing Autism From Other Mental Health Issues NeuropsychiatryDocument11 paginiDifferential Diagnosis and Comorbidity Distinguishing Autism From Other Mental Health Issues NeuropsychiatryJohn Bowen BrownÎncă nu există evaluări
- Management of Recurrent MiscarriagesDocument11 paginiManagement of Recurrent MiscarriagesmarinaÎncă nu există evaluări
- Neuropsych Lec 5 Mental Status ExamDocument2 paginiNeuropsych Lec 5 Mental Status ExamnkivcÎncă nu există evaluări
- DSE Biology Mock (Sunny) Set 1 Paper 1 Section ADocument18 paginiDSE Biology Mock (Sunny) Set 1 Paper 1 Section AwslÎncă nu există evaluări
- Evolution Biology IntroductionDocument22 paginiEvolution Biology IntroductionConstance Viloria100% (1)
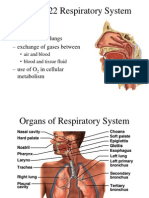
- 22 Respiratory SystemDocument99 pagini22 Respiratory SystemvanderphysÎncă nu există evaluări
- CAR T-Cell Therapy ReviewDocument74 paginiCAR T-Cell Therapy ReviewAviseka AcharyaÎncă nu există evaluări
- Acute Respiratory Distress Syndrome Nursing Management and Interventions - NurseslabsDocument2 paginiAcute Respiratory Distress Syndrome Nursing Management and Interventions - NurseslabsSachin SinghÎncă nu există evaluări
- Test Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner BairDocument35 paginiTest Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner Bairshriekacericg31u3100% (37)
- Antimicrobial Resistance and Clinical Use in Respiratory InfectionsDocument23 paginiAntimicrobial Resistance and Clinical Use in Respiratory InfectionsWamique IzharÎncă nu există evaluări
- Normal Bone Anatomy and PhysiologyDocument9 paginiNormal Bone Anatomy and PhysiologyElsa GonçalvesÎncă nu există evaluări
- Toxicological Evaluation PDFDocument291 paginiToxicological Evaluation PDFMaría SolórzanoÎncă nu există evaluări
- 2nd ICBB ProceedingsDocument117 pagini2nd ICBB ProceedingsLenie Angeles Quiatchon-BaezaÎncă nu există evaluări
- Competent Cell Preparation and TransformationDocument5 paginiCompetent Cell Preparation and TransformationswaroopsanketÎncă nu există evaluări
- DNA Profiling Gizmo Student Exploration SheetDocument5 paginiDNA Profiling Gizmo Student Exploration SheetCarter BonesÎncă nu există evaluări