Documente Academic
Documente Profesional
Documente Cultură
Tema 3. Metodologia Taxonomiei
Încărcat de
macoveidumitruDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Tema 3. Metodologia Taxonomiei
Încărcat de
macoveidumitruDrepturi de autor:
Formate disponibile
Tema 3. 3. METODOLOGIA TAXONOMIEI FILOGENETICE.
Cuvntul filogenie sau filogenez (de la grecescul phylum neam, trib; i genia origine)se utilizeaz n unul din cele dou sensuri pe care le are: 1. Filogenia n sensul larg al cuvntului prezint dezvoltarea istoric a organismelor, ceea ce apropie foarte mult acest termen de termenul evoluie. 2. Filogenia n sensul ngust nu reprezint toate aspectele dezvoltrii istorice, ci numai succesiunea ramificaiilor arborelui filogenetic. n cadrul acestui curs vom vorbi despre filogenie mai mult n sensul ngust al termenului. Este considerat cert faptul, c clasificarea speciilor de organisme trebuie s fie bazat pe principiul ierarhiei stricte: fiecare grup sistematic (taxon) intr n componena unui alt taxon de un nivel mai nalt, iar unul i acelai taxon nu poate face parte din doi sau mai muli taxoni de acelai nivel. Din acest punct de vedere, clasificarea organismelor difer esenial de clasificrile altor elemente naturale (a elementelor i substanelor chimice, de ex.). Din cele mai vechi timpuri sistema de clasificare a organismelor vii a fost creat nu pur i simplu n calitate de clasificare comod, ci ca o reflectare a unei legiti obiective a naturii (cum spunea C. Linnaeus, nu caracterul determin genul, ci genul determin caracterul). Dar numai n rezultatul dezvoltrii teoriei evoluioniste a devenit clar mecanismul biologic, care determin existena unei asemenea legiti. Unitatea unui taxon este determinat de faptul, c toi membrii acestui taxon au provenit de la acelai strmo, iar ierarhia sistemei este determinat de faptul, c mrirea numrului de specii are loc pe calea divergenei (divizarea taxonului parental n dou sau mai multe grupe taxonomice fiice), pe cnd formarea unui taxon din doi sau mai muli taxoni parentali este practic imposibil. Pn n prezent nc nu exist o prere comun, care s reflecte principiile de alctuire a sistemei organismelor vii. Reprezentanii diferitor coli de sistematic declar diferite principii de alctuire a sistemei, iar mai des, principiile declarate nu corespund celor real utilizate
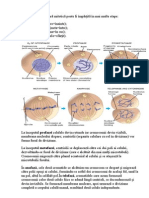
3.1. Metodele de descriere i de reprezentare grafic a filogeniei. Grafic filogenia poate fi reprezentat n form de linie ramificat (adic de dendrogram), care se numete arbore filogenetic, schem filogenetic ori filogram. Filograma poate fi considerat un grafic construit ntr-o manier mai liber dect graficele conform coordonatelor stricte. n cazul prezentrii filogramei n sistemul bidimensional, atunci una dintre dimensiuni este timpul, iar cea dea doua msura deosebirilor dintre organisme. De obicei, dac axa timpului este prezentat vertical, atunci evenimentele timpurii sunt aranjate n parte a de jos a filogramei, iar cele mai trziii n partea superioar; dac axa timpului este orientat orizontal, atunci evenimentele timpurii sunt situate la stnga, iar cele trzii la dreapta. De obicei timpul este reprezentat conform unei anumite scri (mai des scara corespunde erelor i perioadelor geologice, iar uneori este exprimat n mln ani, calculai n baza analizei cu izotopi.)
De mai multe ori ns, scara timpului nu este cunoscut, iar schema este efectuat fr exprimarea timpului. ntr-o astfel de filogram este reprezentat corect doar succesiunea evenimentelor, fr a respecta coordonarea lor cu perioadele concrete de timp i cu durata lor. Paleozoic Mezozoic Cainozoic carbon Permian Triasic Jrasic Cretacic 286 248 213 144 65 Era Perioada 0 Mln ani
Des.1. Arborele filogenetic al AMNIOTA n scara erelor geologice
Des.2. Arborele filogenetic al AMNIOTA fr scara timpului.
De exemplu, conform desenului 2 nu este artat, ce a avut loc mai devreme: apariia psrilor sau separarea mamiferelor n Prototeria i Theria. A doua dimensiune, care pe desenele 1 i 2 este reprezentat vertical arat gradul divirgenelor (deosebirilor).Aici fiecare linie reprezint un taxon existent n timp, care este precutat de ctre cercettori drept un ntreg morfologic. Aceste dou scheme reprezint cu exactitate la nivel de clas i subclas. n cazul, c dorim s exprimm aceast schem cu o exactitate mai mare, atunci n locul fiecrei linii ar trebui s fie desenat un mnunchi de linii sau un tufar de linii ramificate. Uneori, n loc de linii cu aceiai grosime se reprezint dungi, limea crora depinde de numrul speciilor, care formeaz taxonul dat. Distana dintre liniile filogramei reprezint ruptura, care exist ntre formele notate cu linii conform caracterului, care este pus la baza acestei filograme. De obicei, aceste deosebiri morfologie sunt reprezentate fr aplicarea unei scri speciale, cu toate c unii autori tind s atribuie anumite valori numerice deosebirilor, care pot fi calculate prin aplicarea diverselor metode. Orice schem filogenetic nu prezint un desen terminat, ci doar un fragment al unei scheme filogenetice unice a tuturor organismelor , de aceea originea unei filograme separate este prezentat ca o tietur ntr-un loc arbitrar, iar nsi filograma are aspectul unei dendrite nrdcinate. Uneori pentru anumii taxoni este posibil de a reconstrui doar unele legturi filogenetice n interiorul acestui taxon, fr a fi posibil de clarificat n ce mod acest fragment urmeaz a fi conectat la schema filogenetic a organismelor vii. n aa caz avem exemplu de dendrit nenrdcinat.
Reprezentarea grafic a filogenii, asemenea unui arbore se caracterizeaz prin capacitatea sa de a se ramifica. Iat de ce unul din sinonimele acceptate este arborele filogenetic. ns spre deosebire de un arbore real, schema filogenetic nu are un trunchi principal. Pentru schemele filogenetice noiunea de trunchi este echivalent cu cea de ram. Unii cercettori, totui, nominalizeaz drept tulpin acele ramuri, care dau un numr maximal de ramificaii n form de specii. Spre deosebire de arborele adevrat, rdcinile filogramei prezint un punct de nceput al filogramei, selectat arbitrar. Orice arbore filgenetic ntr n componena altuia, mai mare dup complexitate i la rndul su, de obicei este constituit din arbori de ordin mai mic. 3.2. Dou abordri filogenetice Orice organism n mod tradiional este descrs ca unitatea genotipului i fenotipului. Corespunztor, caracterele acestui organism pot fi divizate n caractere genotipice i caractere fenotipice. Valoarea taxonomic a acestor caractere este diferit i cere abordri complet diferite n procesul de selectare i analiz a adatelor. n practica taxonomiei se utilizeaz dou abordri : fenotipic i genotipic. Abordarea fenotipic Abordarea fenotipic este bazat pe analiza fenotipului complexului de caractere realizate ale organismului. Fenotipul, de fapt este obiectul nemijlocit al seleciei naturale. Modificarea lui este determinat de un complex de aciuni din exterior i startegii de rspuns a populaiei la aceste aciuni. Similaritatea strategiilor de rspuns creaz similaritatea fenotipurilor cu caracter convergent, care pot fi obinute n baza diferitor genotipuri. Ca urmare fenotipul nu poate fi un criteriu de ncredere al rudeniei dintre organisme, ci doar un criteriu de similaritate. Cu toate acestea, caracterele, conform crora sunt descrise organismele sunt practic, n exclusivitate fenotipice, iar pe parcursul sutelor de ani se caut metode, care ar permite reconstituirea mersului evolutiv n baza datelor fenotipice. Problema de baz a abordrii fenotipice const n faptul, c fiecare organism posed o mulime enorm de caractere. Utilizarea ntregii sume de caractere este imposibil i neraional din caza volumului enorm de munc, care urmeaz a fi efectuat. Deasemenea aplicarea unui mare numr de caractere secundare, duce la diminuarea valorii unor caractere mai importante. Din aceste considerente, taxonomitii, de regul prefer s opereze cu un mumr restrns de caractere fenotipice considerate mai importante. Esenialitatea caracterului este definit ca greutare taxonomic. Greutatea taxonomic esenialitatea caracterului din punct de vedere al aplicrii lui n clasificarea organismelor vii. Procedura de determinare a greutii taxonomice a caracterului poart denumire de cntrire a caracterelor i poate fi att pur subiectiv, precum i obiectiv din contul aplicrii unor proceduri matematice logice. Abordarea genotipic
Abordarea genotipic este bazat pe analiza genotipului complexului de informaie ereditar a organismului. Genotipul este obiectul indirect al seleciei naturale. Modificarea lui evolutiv are un caracter strict calitativ i este determinat de evenimente ntmpltoare mutaii i recombinaii. Repetarea multipl a unora i acelorai evenimente, precum i apartiia independent a unor genomuri asemntoare, din punct de vedere statistic este puin posibil. Anume din aceste considerente compararea genotipurilor organismelor poate genera informaii veridice despre gradul de rudenie dintre ele, exclude similaritatea convergent i permite de a reconstitui parcursul filogeniei. n aa fel taxonomia genetic este unica metod obiectiv care permite de a reconstrui mersul filogeniei. Implementarea metodelor genetice lrgete esenial posibilitile cercetrilor, transformnd taxonomia ntr-o tiin exact. Dar n acelai timp, descrierea poziiei sistematice absolut corecte a organismului este posibil doar cu condiia descifrrii complete a genomului lui. Pn n prezent acest lucru este realizat pentru un numr mic de specii. Lucrnd cu anumite blocuri de material ereditar, ct de comode n-ar fi acestea pentru realizarea practic, cercettorii sunt pui n condiiile posibilitii de a pierde informaii taxonomice importante. Afar de aceasta, taxonii alctuii n baza datelor genetice sunt caracterizai doar de secvenele nucleotidice, fr a propune i alte caractere diagnostice suplimentare. Aceste caractere diagnostice nucleotidice ale taxonilor sunt greu de aplicat de ctre sistematicieni n lucrrile lor practice. Din aceste motive apare necesitatea fortificrii caracteristicelor genetice cu caractere fenetice adecvate. Transferul orizontal de gene, rspndit n irul organismelor inferioare este unul din fenomenele, care reduce esenial din credibilitatea fa de metodele genetice. Posibilitatea potenial de schimb a blocurilor ntregi de informaie genetic ntre organismele nenrudite reduce la zero convingerea, c succesiunea dat de nucleotide poate fi caracteristic doar formelor nrzdite. Din aceste considerente, atunci cnd datele genetice difer esenial de cele fenotipice, concluzia final despre gradul de rudenie ntre anumii taxoni practic nu poate fi facut fr cercetri suplimentare. Divergenele genetice sunt evidente i cognitive dar n cazul taxonilor superiori. La cercetarea taxonilor inferiori nrudii evidenierea diferenelor este destul de complicat. Astfel, cu toate c taxonomia genetic are o importan fundamental incontestabil necesit confirmare prin date fenetice adecvate. 3.3. Metodele de analiz a datelor taxonomice Cercetrile taxonomice demult au depit limitele combinrii arbitrare a taxonilor n conformitate cu preferinele ori viziunile cercettorilor. n ultimele decenii a fost creat o baz metodologic puternic, care permite n cea mai mare msur de a standardiza i a face ct mai obiectiv activitatea taxonomitilor. Cercetrile contemporane n domeniul sistematicii implic utilizarea anumitor metode speciale. Metoda tipologic
Metoda tipologic a fost fondat n baza analizei caracterelor unui organism. De fapt este o analiz cladistic a caracterelor fenotipice , care s-a stabilit n secolul XVIII. Construirea sistemei n baza metodei tipologice include urmtoarele etape: 1. Toi indivizii sunt repartizai n grupuri n dependen de aceea, cu care dintre tipurile (etaloanele) cunoscute sunt identici acetea. De cele mai multe ori se utilizeaz nu etaloanele ci descrierea lor n surcele bibliografice. 2. Se efectueaz evidena caracterelor grupurilor create. 3. Se efectueaz evaluarea esenialitii caracterelor evideniate din punct de vedere al clasificrii cntrirea caracterelor. n cadrul metodei tipologice aceast procedur poart un caracter pur subiectiv li se bazeaz pe concepia cercettorului despre evoluia grupului cercetat. n rezultatul evalurii fiecrui caracter i este atribuit o anumit greutate taxonomic. 4. n dependen de greutatea taxonomic acordat , caracterele sunt repartizate n grupuri, iar fiecrui grup i este acordat un anumit rang taxonomic. De exemplu, pentru alge caracterul componenei pigmenilor n cloroplaste caracterizeaz Filumurile, morfologia talului clasele, structura aparatului flagelar ordinele, iar tipul nveliurilor celulare familiile. Asrfel, analiznd caracterelor unui exemplar, care aparine unei anumite specii, cercettorul recunoate, c nu toate caracterele sunt tipice taxonului dat la nivel de specie. Fiecare exemplar poart caractere de gen, familie, ordin, sau din contra, carctere de varietate, form. Comparnd ntre ele speciile, cercettorul opereaz cu caractere strict specifice. Dac aceste specii au fors atribuite unui gen, atunci urmeaz a lucra cu caractere de gen. La atribuirea exemplarului la o familie, n lucru se vor lua caracterele de familie .a.m.d. Asemenea complexe de caractere, care permit deosebirea unui taxon de altul de acelai rang poart numele de grupuri tip. 5. Ultima etap a analizei tipologice const n delimitarea taxonilor. Pe parcursul acestui procespas cu pas, n corespundere cu greutatea taxonomic se iau n considerare strile caracterelor fiecrei grupe. La nceput sunt cercetate caracterele cu o greutate taxonomic mai mic, iar organismele sunt repartizate n taxoni inferiori. Dup aceasta sunt stuate caracterele cu o greutate taxonomic mai mare, iar taxonii mici sunt unii n taxoni superiori. Ca rezultat se alctuiete o sistem clar a grupului studiat. Avantaje: Metoda tipologic se caracterizeaz printr-o istorie lung i tradiii adnci. A adus n sistematic multiple idei constructive, aa ca delimitarea caracterilor de strile lor; noiunea de grup tip; precum i noiunea de exemplar tip, pe care se bazeaz practic toat nomenclatura biologic.Sistemele alctuite n baza metodei tipologice se caracterizeaz prin caracter logic, comoditate n aplicare, deoarece sunt cel mai des alctuite dup principiul cheii de determinator. La clasificarea organismelor microscopice puin studiate metoda tipologic este indicat. Neajunsuri
Principalul neajuns al acestei metode este subiectivitatea principial a aprecierii valorii caracterelor. Este practic imposibil de a stabili nite principii clare, universale, fundamentate de evaluare a caracterelor. Astfel, de pe poziii teoretice metoda tipologic este foarte vulnerabil, fiind n realitate nu att o metod tiinific, ct o sistem de reguli, utile n lucru de rutin al taxonomitilor.
Metoda sistematico-evoluionist La nceputul anilor 1950 teoria taxonomic a fost supus aciunii unui nou curent sistematica evoluionist. Fondatorii acestui curent sunt ornitologul Mayer i paleontologul Simpson, studiind diversitatea vertebratelor, au ajuns la concluzia , c este absolut imposibil de a descrie aceast diversitate cu ajutorul metodelor tradiionale i a principiilor unificate. Spre deosebire de adepii metodei tipologice, ei afirmau, c caracterul nu poate avea o importan universal: n clasificarea unui grup un caracter paote fi important, iar n cazul unui alt grup urmeaz a fi ignoratAstfel de caracter este si rudenia dintre taxonii superiori E.Mayr a elaborat abordarea populaional a acestei metode. Conform concepiei lui, cercetarea taxonomic trebuie s fie bazat nu pe studierea exemplarelor tip, ci pe compararea populaiilor i aprecierea variabilitii intraspecifice. J.Simpson a expus aspectul paleontologic al sistematicii evoluioniste. Conform opiniei lui, stabilirea filogenezei unui anumit grup este un lucru util, dar nu este scopul de baz al sistematicii. Scopul sistematicii const n a nelege n ce mod sunt legate i grupate organismele din punct de vedere biologic. Simpson considera c o sistem biologic trebuie s conin ct mai mult informaie. Clasificarea i filogenia sunt lucruri diferite, dar legate ntre ele. Sistematica evoluionist clasic tinde s includ ct mai mult informaie de natur evoluionist n sistema lumii organice. Clasificarea formal este o ncercare de a reflecta n acelai timp legturile de rudenie i gradul de divergen ntre organismele nrudite. E clar, c o corespundere absolut ntre clasificare i filogenie este imposibil. n baza acestor idei a fost elaborat o sistem original de analiz a datelor taxonomice, care const din trei etape de baz. 1. Se efectuiaz o cercetare multilateral a unui grup de organisme conform tuturor parametrilor fenotipici: morfologici, fiziologici, ecologici, etologici .a. n limita posibilitilor, studiul include i cercetarea populaiilor reale, a structurii lor, hotarelor arealului, gradului de izolare .a. Se cerceteaz deasemenea filogeneza grupului, preferenial conform datelor paleontologice. 2. Din complexul de date, cercettorul n fiecare caz n parte le selecteaz pe acele, care au o valoare mai mare.Asemnarea dintre organisme i rudenia presupus dintre acestea se precaut n calitate de parametri echivaleni , iar cercettorul are dreptul s decid, n ce msur relaiile de rudenie va influena strucrura clasificaiei. Cu alte cuvinte are loc aceiai apreciere a valorii caracterelor, dar de data aceasta cu participarea unui nou parametru filogenia. 3. n baza criteriilor selectate, organismele se grupeaz n taxoni. Dac n limitele unui taxon superior (de exemplu clasa A) se evideniaz un grup (ordinul b), care a obinut pe parcursul evoluiei anumite caractere distinctive fa de alte ordine ale acestei clase ,
taxonomistul poate separa acest ordin n calitate de clas separat clasa B. n acest caz lasa A este considerat strmosul actual al clasei B. Un asemenea raport al taxonilor, cunoscut ca parafilie (termenul va fi explicat n continuare) este recunoscut dar de adepii metodei sistematico-evoluioniste. Avantaje: Cercetrile fundamentale ale adepilor metodei sistematico-evoluioniste au contribuit esenial la studiu organismelor superioare. Ei au demonstart caraterul limitat al clasificrilor tipologice i au luat asupra sa efortul titanic de a studia fiecare specie. n stusiul taxonilor inferiori ai organismelor superioare aceast metod este o direcie prioritar n continuare. Neajunsuri: Volumul mare de lucru implicat n realizarea metodei sistematico-evoluioniste face posibil aplicarea ei doar pentru taxonii inferiori. n cazul taxonilor superiori eficiena metodei este limitat de imposibilitatea aplicrii criteriilor reproductiv i ecologo-populaionale.n prezent nu exist metode, care permit de a aprecia caracterele grupurilor descendente i de a aprecia, care dintre ele urmeaz a fi scoase dup limitele grupului strmo, i care urmeaz s fie lsate n componena lui. Deaceea recunoaterea grupelor parafietice duce dup sine subiectivizarea cercetrilor taxonomice. Metoda nu poate fi aplicat atunci, cnd lipsete posibilitatea de studiu a populaiei i efecturii ncrucirilor: lucru cu microorganismele n materiale naturale, cercetrile n ierbare i colecii, sau atunci cnd obiectul studiat nu este capabil de a se reproduce pe cale sexuat. Ca urmare, aplicarea acestei metode n domeniul micologiei, algologiei, protozoologiei, bacteriologiei este limitat. Metoda fenetic Spre deosebire de alte principii de clasificare ale organismelor, fenetica nu ia n consideraie filogenia organismelor studiate. La baza feneticii se afl tendina de a transforma sistematica ntr-un domeniu ct mai obiectiv, empiric, independent de viziuni subiective i diverse speculaii. n aceast ordine de idei filogenia este considerat ca inutilizabil pentru sistematica obiectiv deoarece constituie o ipotez, care nu poate fi demonstrat obiectiv i conine premise subiective. ntr-adevr, filogenia nu poate fi studiat prin metode directe, i pentru reconstrucia filogenetic este nevoie de a utiliza metode indirecte, care nu ntotdeauna dau un rezultat univoc. Astfel, conform opiniei adepilor feneticii, baza adecvat a sistematicii n loc de filogenie trebuie s fie constituit din caracterele organismelor, deoarece caracterele exist obiectiv i nu sunt ipotetice. Se consider, c toat diversitatea caracterelor organismelor const din uniti ale fenotipului fene (numite astfel prin analogie cu genele, deoarece se considera, c fiecare fen este determinat de existena unei gene unitate elementar a genotipului). n fenetic toate fenele se consider echivalente, deoarece, selectarea fenelor mai mult sau mai puin importante este subiectiv i ar duce la o clasificare subiectiv. Idea a fost formulat de ctre Adanson. Este evident, c dac la alctuirea unei sisteme vor fi folosite caracterele, toate fiind considerate echivalente, atunci o asemenea sistem nu va avea caracter ierarhic, deoarece unele caractere se ntlnesc n diferite combinaii. De asemenea sistema alctuit nu va fi univoc, deoarece va depinde de caracterele luate n consideraie la alctuirea ei. n acelai timp fenetica indic asupra strictei necesiti de a alctui o sistem strict ierarhic i univoc. Pentru atingerea acestui obiectiv n fenetic se utilizeaz metodele matematice (analiza
cluster), care sunt bazate pe calcularea indicelui mediu de asemnare n baza repartizrii multiple a caracterelor. Reprezentarea grafic a rezultatului unei asemenea analize prezint un arbore ierarhic unic ramificat (dendrogram), care poart numele de fenogram. Fenograma nu trebuie confundat cu filograma (arborele filogenetic). ntr-o filogram ramificaiile indic asupra gradului de nrudire a grupelor filogenetice, pe cnd n fenogram fiecare ramificaie reflect nivelul de asemnare calculat pentru un anumit grup arbitrar de caractere, selectate arbitrar prin metodele matematice. Din aceste considerente, pentru o anumit grup de organisme exist o singur filogram corect, n timp ce pentru aceiai grup de organisme exist un numr nelimitat de fenograme corecte. Organismele, care se caracterizeaz printr-un anumit nivel de asemnare se unesc ntr-un taxon , numit fenon . n aa mod un fenon ntrunete organismele dup nivelul de asemnare, independent de gradul lor de rudenie. n dependen de caracterele selectate pentru analiz i de metodele matematice utilizate se pot obine clasificri fenetice diverse pentru unele i aceleai organisme. Se vehicula idea, c cu ct mai multe caractere vor fi luate n consideraie n procesul de analiz fenetic i ct mai performante vor fi metodele matematice aplicare, cu att diferitele clasificri fenetice ale unora i acelorai organisme se vor apropia spre o clasificare ideal natural. ns, deoarece filogenia este ignorat contient de ctre analiza fenetic, este neclar, n baza cror mecanisme naturale ar fi posibil acest lucru. Experiena analizei matematice a demonstrat, c la mrirea numrului de caractere incluse n calcul tot una nu are loc apropierea fenogramelor.. n aa mod, fenetica nu numai c nu salveaz sistematica de ipoteze, ci din contra, o pune n dependen de ipoteza vag a existenei unei sisteme naturale, nebazate pe filogenie.fenetica nu numai c nu diminuiaz elementul subiectiv n sistematic, ci din contra, mrete subiectivismul clasificrii, deoarece presupune selectarea subiectiv att a caracterelor, ct i a metodelor matematice de analiz. Conform caracterelor Conform caracterelor femelei mature cariotipice
Fig. 4.3. Diferite fenograme pentru acelai grup de specii din genul Chironomus obinute n baza analizei diferitor caractere
Metoda elaborat iniial pe baza acestor principii a fost numit taxonomie numeric n anul 1965 Mayr a propus denumirea acestei direcii ca FENETIC. Deasemenea aceat direcie mai este numit i taxonomie Adanson. Pe msura dezvoltrii matematicii statistice au fost elaborate programe de compiuter care au permis o rspndire larg a metodei fenetice.Cea mai mare popularitate printre ele au obinut-o Excel, Statistica. PAUP. Datorit acestor programe practic cele mai complicate cercetri numerice sunt acum accesibile fiecrui sistematician. Construirea clasificrii cu ajutorul metodei fenetice include urmtoarele etape: 1. La prima etap se efectuaiz descrierea detaliat a unui numr maximal posibil de caractere calitative i cantitative ale organismului. Este necesar de a avea rezultatele multiplelor msurri (nregistrri) ale caracterului . Pentru caracterele cantitative se utilizeaz rezultatele msurrilor, iar pentru caracterele calitative se aplic cifrarea lor n form de cod numeric. n cel mai simplu caz se utilizaz codul 0/1, adic echivalentul lipsete/este. n caz c caracterul calitativ poate avea mai multe variate alternatve, de exemplu culoarea poate fi gri, neagr, maro, atunci caracterele pot fi codate astfel gri=1; maro=2; negru=3. Toate datele numerice obinute n aa mod se ntroduc ntr-un tabel unic tabelul primar. Completarea acestui tabel se face fr aplicarea posibilitilor programelor compiuterizate i este etapa, ce implic cel mai mare volum de lucru. 2. Datele colectate sunt supuse analizei statistice cu utilizarea programei compiuterizate. Se clarfic crui tip de repartizare matematic se supune fiecare ir al fiecrui caracter, se calculeaz media valorilor obinute, devierea standart i ali parametri statistici. La aceast etap este recomandat de a efectua analiza corelaional a caracterelor, care permite de a evidenia n ce msur aceste manifestri se aseamn la diferii taxoni. De exemplu caracterele 1 i 2 pot fi considerate ca corelante fa de taxonii A i B dac n taxonul A ambele caractere se abfl n starea 0 , iar la taxonul B n starea 1. 3. Toate datele matricii primare se supun standartizate (normalizate) are loc recodificarea tuturor caracterelor n conformitate cu o scar unic de la -1 la +1. Aceasta permite de a trece toate valorile din matrice ntr-o form comparabil. Standardizarea se face n conformitate cu formula Xn=(X-X)/2D, unde Xn este valoarea standardizat, X valoarea iniial, X valoarea medie a parametrului, iar D dispersia parametrului X. n rezultat se creaz o matrice secundar. Matricea primar Taxon Culoarea A 0 B 0 C 1 D 1 Diametrul 1 1 2 5 nlimea 5 10 12 10 Limea 1 1 2 3 Matricea secundar Taxon Culoarea A -0,9 B -0,9 C 0,9 D 0,9 Diametrul -0,7 -0,7 -0,1 1,5 nlimea -1,4 0,3 0,9 0,9 Limea -0,8 -0,8 1,3 1,3
4. Valorile din matricea secundar se analizeaz de pe poziia asemnrilor i deosebirilor. Exist dou procedee de baz care permit de a realiza aceast analiz: analiza de cluster i analiza factorial. n cazul analizei de cluster toate valorile care sunt introduse n matrice se grupeaz n perechi n toate combinaiile posibile, iar pentru fiecare pereche se calculeaz msura
asemnrii matematice, de exemplu, distana Euklidian (distana liniar ntre punctele taxoni n sistema de coordoate a dou caractere). n cazul analizei factoriale compiuterul analizeaz concomitent toi taxonii n spaiul multidimensional al caracterelor i selecteaz 2-3 factori ( de regul, care nu corespund caracterelor concrete), care permit de a exprima mai deplin distana dintre taxoni n n spaiul bi- sau tridimensional corespunztor. 5. Compiuterul genereaz o reprezentare grafic a datelor, obinute n rezultatul analizei asemnrilor i deosebirilor. Datele analizei de cluster sunt de obicei reprezentate n form de diagram de cluster (dendrogram, fenogram) schem n form de arbore, n care distana dintre taxoni este reprezentat printr-o linie de legtur. Rezultatele analizei factoriale sunt prezentate n form de sistemde coordonate bi- sau tridimensionale. n care taxonii sunt artai sub form de puncte. 6. Taxonii sunt grupai dup principiul apropierii maxime n msurri, care sunt propuse de ctre analiza de cluster ori de cea factorial. Grupele obinute, la rndul lor sunt grupate n grupiri de un ordin mai nalt, pn la obinerea unui singur supertaxon, care i include pe toi ceilali. Fiecrui grup format i se atribuie un anumit rang taxonomic. Ca rezultat seobine o sistem ierarhic. Avantaje: Avantajul metodei const n caracterul ei obiectiv i stricteea tiinific. Aparatul matematic puternic permite de a aprecia exact asemnarea i deosebirea multifactorial ntre diferite organisme, de a depista hiaturile (rupturile) ntre grupurile nrudite, de a evalua gradul de corelare ntre caracterele analizate. Acest lucru este necesar n special n procesul de studiu al taxonilor inferiori ( genuri, specii, varieti) i formelor fosile. Dezavantaje: Egalitatea caracterelor este o presupunere foarte dubioas. Independent de abordare, diferite caractere vor avea o importan diferit n alctuirea sistemei. Clasificrile fenetice, care nu iau n veder acest lucru, de obicei se dovedesc a fi teoretic lipsite de coninut. Analiza numeric este bazat pe evaluarea asemnrii dintre organisme i nu poate depista nivelul de nrudire a lor, deoarece asemnrile des au un caracter convergent.Cu toate c diagrama de cluster are aspectul unui arbore filogenetic, de fapt nu are nimic comun cu acesta, deoarece nu descrie mersul evoluiei, deaceea are o utilizare foarte nensemnat n analiza filogenetic. Deoarece siatematica contemporan se bazeaz n primul rnd pe abordarea filogenetic, utilizarea metodei numerice devine neactual, n special n ceea ce privete taxonii superiori. Metoda cladistic Fondatorul analizei cladistice (de la grecescul cladus ram) este entomologul german Willy Henning (1913-1976), care n anul 1950 public cartea "Grundzuge einer Theorie der phylogenetischen Systematik" (traducerea n englez "Phylogenetic systematics", care l-a fcut de fapt celebru a aprut n SUA n 1966). De fapt n lucrarea sa Henning nu a propus o metod nou. El a utilizat metodele, pe care biologii le foloseau de foarte mult timp (chiar i Lamarck n Filozofia zoologiei reproduce o schem filogenetic, care putea fi construit numai prin metode de cladistic). Meritul lui Henning const n aceea, c el pentru prima dat a descris succesiv metodele de reconstrucie filogenetic, iar lucrarea sa i-a impus pe ali cercettori s formuleze clar acele principii, pe care le folosesc n procesul reconstruciei filogenetice. Meritul lui Henning mai const i n introducerea unor termeni foarte importani.
Termenii apomorfia i plesiomorfia Termenul de apomorfie (grecescul apo de la) sau caracter apomorf nseamn caracter progresiv, secundar, evolutiv. Termenul de plesiomorfie (grecescul plesios apropiat) sau caracter plesiomorf nseamn caracter primitiv, iniial, ancestral, de la strmo, sau caracter generalizat. Apomorfia i plesiomorfia sunt dou stri diferite ale caracterului pn la i dup modificarea creia a fost supus evolutiv acest caracter. Aceste noiuni de asemenea sunt relative. Dac evoluia merge de la starea A a caracterului spre starea B, apoi spre C, atunci Starea B este apomorf fa de A i plesiomorf fa de C. Termenii dai au un singur sens direcia modificrii evolutive a caracterului dat n cazul dat. Apomorfia, care deosebete un taxon de toi ceilali taxoni se numete autoapomorfie, iar cea, care este caracteristic pentru mai muli taxoni se numete sinapomorfie. Plesiomorfia comun pentru mai muli taxoni se numete simplesiomorfie. Analiza filogenetic pe baza apomorfiilor Analiza cladistic se bazeaz pe presupunerea, c modificrile evoluioniste sunt ireversibile. Deci, dac mai muli taxoni au acelai caracter apomorf (sinapomorfie), atunci aceti taxoni au motenit caracterul dat de la un strmo comun, i deci, aceste grupe taxonomice formeaz un ram filogenetic comun. Pentru a demonstra c civa taxoni formeaz un ram filogenetic comun trebuie de demonstrat prezena a cel puin unei sinapomorfii, care este o autoapomorfie pentru toi taxonii precutai, ceea ce demonstreaz, c aceti taxoni au motenit caracterul progresiv corespunztor de la acelai strmo i formeaz o singur ramur filogenetic. Astfel, pe desenul 3 apomorfia caracterului 3 este o sinapomorfie pentru taxonii C i D i o autoapomorfie pentru ramura C-D. Aceast apomorfie demonstreaz relaia existen a ramurii C-D membrii creia au provenit de la un strmo ipotetic comun e, la care a i aprut apomorfia acestui caracter. n acelai timp prezena simplesiomorfiilor la civa taxoni nu vorbete despre aceea, c acetea au un strmo comun. Astfel plesiomorfia caracterului 3 la taxonii A i B nu ne permite s vorbim despre originea lor comun sau rudenia lor. Putem spune doar c aceti doi taxoni nu aparin ramurii filogenetice C-D.Simplesiomorfia caracterului 2 la taxonii B,C,D de asemenea nu ne permite s afirmm apartenena lor la aceiai ramur filogenetic, n schimb putem afirma acest lucru n baza siapomorfiei caracterului 1.
Des.3. Reconstrucia relaiilor filogenetice dintre 4 taxoni (A,B,C,D) n baza analizei a ase caractere (1-6); e i f - taxoni strmoi ipotetici; dreptunghiuri necolorate stare plesiomorf a caracterului; dreptunghiuri negre starea apomorf a caracterelor; linia gri sinapomorfie. Astfel, arborele filogenetic alctuit doar n baza analizei cladistice poart numele de cladogram i se deosebete de alte tipuri de arbori filogenetici prin aceea, c conine informaie doar referitor la succesiunea ramificaiilor, fr referin la timpul absolut cnd are loc ramificarea i la gradul de divergen ntre ramuri. Determinarea polaritii caracterului Pentru determinarea polaritii caracterului (adic clarificarea strii caracterului plesiomorf ori apomorf) se aplic principiul grupului extern. Pentru a exemplifica acest principiu ne vom ntoarce la des. 3. Am clarificat deja existena unor ramuri filogenetice, de exemplu ramura B-D. pentru a clarifica relaiile n interiorul acestui grup se cerceteaz caracterele, care se deosebesc la reprezentanii din diferii taxoni ai grupului (n cazul nostru este vorba despre caracterele 3-6). Starea plesiomorf a caracterului este acea, care n afar de aceast grup se mai ntlnete n alte grupuri, care ocup o poziie extern fa de taxonii B-D. n acelai timp starea apomorf a caracterului este acea stare, care n afara grupului dat nu se ntlnete la alte grupuri externe. De exemplu printre Lepidoptera sunt foarte muli reprezentani, aparatul bucal al crora este de tip de supt, format dintr-o tromp rsucit i 2 palpi. n acelai timp se mai ntlnete o familie Micropterygide, la care aparatul de supt are o structur complicat fiind combinat cu cel de ros. Ar fi de prisos i nerezultativ s discutm despre aceea, care tip al aparatului bucal este plesiomorf i care este apomorf, ce este mai important i n ce direcie a mers procesul evolutiv. Deoarece aici avem dea face cu direcia de aciune a seleciei naturale, care se poate manifesta foarte diferit n diferite condiii. De aceea unica ieire este de a compara aceste tipuri de structur a aparatului bucal att n interiorul grupei, precum i n afara ei. A fost demonstart, c apart de tipul trompei simple nu se ntlnete dect la reprezentanii ordinului Lepidoptera, pe cnd aparat bucal de tip de ros este larg
rspndit n alte grupe sistematice. Astfel, aparatul bucal de tip ca la Micropterygide este un caracter de tip plesiomorf, pe cnd aparatul de tip de supt este un caracter apomorf. Noiune de mono-; poli-; para- i holofilie n dependen de natura filogenetic a taxonilor, acetia se mpart n monofiletici (cu variaiile parafiletici i holofiletici) i polifiletici. Termenii de monofilie i polifilie au fost ntrodui pentru prima dat de ctre Haeckel n sec. XIX i se utilizeaz de atunci, nectnd la faptul, c nu au fost caracterizai prin definiii stricte. Din cauza lipsei unei definiii clare muli cercettori au propus diferite variante de interpretare a termenilor, uneori chiar contradictorii. Cel mai general mod de exprimare a termenilor dai este urmtorul: taxon monofiletic este taxonul care provine de la un strmo, iar taxon polifiletic este cel, care provine de la mai muli strmoi diferii. Dar, pornind de la faptul, c toate organismele au provenit de la un strmo comun, atunci o asemenea determinare a definiiei este evident insuficient, deoarece conform ei orice taxon pn la urm este monofiletic. Una din definiiile de alternativ propus de Mayr, 1971, sun n modul urmtor: taxonul monofiletic este un asemenea taxon , care provine pe una sau mai multe linii de la un taxon nemijlocit parental de acelai nivel, sau de un nivel mai mic. Aceast definiie este criticat pe larg, deoarece este n dependen de o noiune artificial, aa ca nivelul taxonului. n aceast ordine de idei ar fi posibil de a numi drept monofiletic orice taxon, dac determinm ntr-un mod care corespunde cazului nivelul lui. Conform definiiei acceptate n prezent de ctre sistematicienii diferitor coli taxon MONOFILETIC n sensul larg al cuvntului se numete acela, care include n sine strmoul, comun pentru toi membrii acestui taxon, precum i toate ramurile filogenetice, care duc de la acest strmo la fiecare membru al taxonului. Deoarece strmoul taxonului, precum i alte forme parentale de obicei lipsesc din natura contemporan i sunt necunoscute cercettorilor, este mai exact de a spune, c strmoul nu este parte component a taxonului monofiletic, ci corespunde diagnosticului taxonului dat. Taxonii monofiletici n sensul larg al cuvntului se mpart n holofiletice (adic monofiletici n sensul ngust) i parafiletici. Taxon holofiletic sau strict monofiletic se numete acela, care pe lng faptul, c include n sine strmoul, i toate ramurile filogenetice, care duc la fiecare membru al taxonului, mai include i toi descendenii strmoului comun. Pe desenul 4. holofiletice sunt taxoanele A-B-C-D-E i B-CD. Printre caracterele, care caracterizeaz taxonul holofiletic se numr de obicei apomorfiile, sau caracterele constitutive ale taxonului, care prezint sinapomorfii pentru toate grupele, care intr n componena taxonului. Astfel, apomorfia aprut la strmoul comun al taxonului holofiletic se va pstra la toi descendenii acestui strmo, iar deoarece, descendenii acestui strmo nu pot fi gsii n afara taxonului dat, nici apomorfia dat nu poate fi gsit dect n acest taxon holofiletic. Din aceast definiie rezult, c taxonul holofiletic nu poare fi taxon strmo pentru un alt taxon, deoarece, toate formele, care provin de la membrii taxonului holofiletic sunt inclui anume n acest taxon. Dac taxonul polifiletic este separat printr-o curb nchis pe arborele filogenetic, atunci aceast curb va ntretia arborele filogenetic doar ntr-un singur punct, care corespunde intrrii n taxon. Prima parte a termenului holofilie provine de la grecescul holos, care nseamn ntreg. Astfel,
un taxon holofiletic este constituit dintr-o ramur filogenetic ntreag, care este conectat la arborele filogenetic doar ntr-un singur punct baza acestei ramuri, i include toate ramificaiile ei. Taxon parafiletic se numete acela, care include n sine strmoul, toate ramurile filogenetice, care duc la fiecare membru al taxonului, dar nu include toi descendenii strmoului comun. Spre deosebire de taxonul holofiletic, taxonul parafiletic este strmoul unui alt taxon (sau mai multor taxoni) i se deosebete de acesta (acetia) doar prin plesiomorfii. Pe desenul 4 taxonul parafiletic A-B-D-E este taxon strmo pentru taxonul C. Dac pe schema filogenetic taxonul parafiletic este ncercuit cu o curb nchis, atunci aceasta va intersecta schema n mai multe puncte o dat la intrarea n taxon i o dat sau de mai multe ori la ieirea din el. Taxon POLIFILETIC este acela, care nu include strmoul comun pentru toi membrii taxonului, ori care nu include anumite segmente ale ramurilor filogenetice, care unesc strmoul comun cu careva din membrii taxonului. Cu alte cuvinte strmoii nu corespund diagnosticului de taxon. Caracterele, conform crora se face diagnosticul taxonului au aprut pe parcursul evoluiei n mod independent de mai multe ori. Pe figura 4 sunt artai taxonii polifiletici A-B i A-B-E. dac taxonul polifiletic este ncercuit pe arborele filogenetic printr-o curb nchis, atunci aceast curb va intersecta n mai multe puncte arborele de cel puin 2 ori la diferite intrri n taxon. Pentru a evita situaiile confuze, de obicei termenul de monofilie este nlocuit cu varianta corespunztoare de holofilie i parafilie, fiecare dintre care este determinat destul de univoc.
a) Holofilie (a)
b) Parafilie(b)
c)
d) Polifilie(c,d)
Des.1. Taxoni holofiletici (a), parafiletici (b) i polifiletici (c i d) Exemplu Exemplul clasic de taxon parafiletic este clasa Reptilia. Conform teoriei filogeniei acestei grupe, de la dinozauri au provenit psrile. Dinozaurii sunt parte component a taxonului Reptilia, pe cnd Aves nu. Pentru psri este separat un alt taxon clasa psrilor. Clasa Aves este un taxon holofiletic i se caracterizeaz printr-un complex de apomorfii, inclusiv prezena penelor, formaiuni pieloase, provenite din modificarea solzilor la reptile. n acelai timp, taxonul parafiletic Reptilia nu are i nu poate avea apomorfii caracteristice clasei Aves, iar toate caracterele, care deosebesc clasa reptile de clasa psri sunt caractere plesiomorfe (de exemplu prezena solzilor). Fig. 5 .
Fig.5. Taxonul parafiletic Reptilia.
Tabelul 1. Noiuni de Holofilie, Parafilie i Polifilie Taxonul studiat Include toi descendenii Include unii descendeni ai strmoului comun strmoului comun Include strmoul comun Holofiletic(sau strict Parafiletic monofiletic) Nu include strmoul comun polifiletic
Domenii de aplicare a analizei cladistice. Analiza cladistic n forma sa clasic este destinat reconstruciei filogenetice, care se bazeaz pe divergene, cnd schema filogenetic are aspect de arbore ascendent ierarhic, i nu de reea. Analiza cladistic nu este destinat studiul acelor cazuri de evoluie, cnd la mecanismele obinuite de divergen se altur asemenea factori ca hibridizarea, simbiogeneza sau transducia; n aceste cazuri arborele filogenetic capt aspect de reea. n linii generale, analiza cladistic este aplicat pentru analiza taxonilor nu mai mici dect specia. ntre taxonii intraspecifici nu exist izolare reproductiv i deci, relaiile de rudenie dintre ele nu poart un caracter de arbore filogenetic, ci de reea, de aceea nu pot fi depistate prin analiz cladistic. Muli cercettori aplic analiza cladistic pentru stabilirea legturilor filogenetice ntre specii. n linii generale aceasta este posibil, dar procentul de erori este destul de mare din cauza c afar de sinapomorfii (caractere motenite de la acelai strmo) speciile apropiate pot manifesta diferite caractere omoloage, aprute independent. Aceasta se ntmpl, deoarece foarte des se motenete nu caracterul fenotipic, ci gena, sau combinaia de gene, care n anumite circumstane asigur ori nu exprimarea caracterului dat. Caracterul omolog, care apare independent poate fi confundant uor cu caracterul sinapomorf, care se manifest doar n anumite circumstane. n acest caz analiza cladistic va duce la rezultate eronate. Cu ct mai apropiate din punct de vedere filogenetic sunt taxonii supui analizei cladistice, cu att mai mare este probabilitii apariiei erorilor, deoarece n aceste grupe este foarte mare probabilitatea apariiei caracterelor omoloage independente. Dac fiecare din taxonii supui analizei cladistice conine un numr mare de specii diverse, este de ateptat c caracterele ascunse comune ale taxonului se vor manifesta la unele din speciile lui. n acest caz asemenea caractere se exclud din analiza cladistic. Iat de ce, cu ct mai nalt este gradul taxonului examinat i cu ct mai multe grupe taxonomice incluse cuprinde el, cu att
rezultatele obinute sunt mai aproape de adevr. Cele mai puin autentice rezultate se obin la analiza cladistic a speciilor. Aplicarea caracterelor moleculare n analiza cladistic Aplicarea caracterelor moleculare ale taxonilor asigur o depistare mai eficient a legturilor filogenetice ntre ele. De obicei se efectuiaz analiza cladistic n baza succesiunii aminoacizilor n componena proteinelor precum i succesiunea nucleotidelor n anumite molecule de ADN i ARN ale reprezentanilor diferitor taxoni. La prima vedere s-ar prea, c la aplicarea analizei cladistice a succesiunii bazelor azotate rezultatul obinut va fi unic pentru reconstrucia filogenetic. Pentru analiza caracterelor moleculare se utilizeaz soft-ri specializate, care se bazeaz pe presupunerea, c toate mutaiile sunt equiprobabile. n aa caz apariia unei mutaii de acelai tip n unul i acelai loc la diferii taxoni este puin probabil. Ca urmare se presupune c prezena unei mutaii similare la diferii taxoni vorbete despre sinapomorfia lor i despre faptul, c a fost motenit de la un strmo comun, ceea ce permite stabilirea legturilor filogenetice ntre taxoni. n realitate ns lucrurile stau altfel i mutaiile nu sunt equiprobabile. Genomul organismelor este nzestrat cu mecanisme complicate de reglare, iar n componena materialului genetic sunt structuri cu capacitate foarte nalt de mutaie i invers, cu capacitate de mutaie nul. Sinapomorfii credibile sunt acelea, care apar n segmentele caracterizate prin activitate mutaional joas i foarte joas. Deacea, pentru reuita analizei cladistice n baza caracterelor moleculare este foarte important selctarea segmentelor, compararea crora ne va oferi informaie satisfctoare pentru stabilirea legturilor filogenetice. Din considerente c stabilirea succesiunii nucleotidelor este un proces foarte laborios, des este suficient de a efectua examenul comparat al unor pri ale genotipului. O metod, care permite compararea genitipurilor integre este metoda hibridrii moleculare ADN. ADN-ul colectat de la 2 taxoni diferii supui comparrii se supune despiralizrii cu obinerea fragmentelor monocatenare i se amestec. Prin crearea condiiilor speciale se provoac formarea spiralelor bicatenare complementare . Separnd segmentele bicatenare de cele monocatenare se calculeaz partea regiunilor complementare adic a regiunilor identice pentru ambii taxoni. Pentru a compara mai mult dect doi taxoni unul din ei este selectat n calitate de reper i se amestec consecutiv ADN-ul lui cu ADN celorlali taxoni (cu cte unul). Rezultatul analizei comparative nu este influenat de faptul, care dintre taxoni a fost selectat drept reper. n cazul unui asemenea tip de analiz caracterele comune (secvenele nucleotidice identice) nu pot fi separate n sinapomorfe i sinplesiomorfe, deacea nu poate fi aplicat analiza cladistic. Hibridizarea molecular a ADN permite stabilirea exact a gradului de similitudine genetic ntre oricare specii, dar nu permite stabilirea legturilor filogenetice ntre ele.
Principiile cladisticii: Analiza cladistic este bazat pe un complex de principii teoretice, cunoaterea crora permite de a aprecia obiectiv posibilitile acestui rip de analiz. Exist patru principii de baz ale cladisticii:
Principiul siapomorfiei; Rincipiul monofiliei stricte; Principiul grupurilor nfrite; Principiul parsimoniei.
Principiul Sinapomorfiei Toate caracterele unui organism sunt apreciate de ctre un cladist din punct de vedere al naturii lui primitive (pleziomorfiei) sau progresive (apomorfiei). Dac civa taxoni sunt unii printrun caracter apomorf, atunci se vorbete despre sinapomorfii. Apariia la doi taxoni a unui caracter plesiomorf se numete sinpleyiomorfie. n cele din urm, prezena la un taxon a unor caractere unicale, care nu se ntlnesc la ali taxoni poart denumirea de autapomorfie. Esena principiului sinapomorfiei const n aceea c pentru construirea clasificaiei filogenetice au importan numai caracterele sinapomorfe. Caracterele pleziomorfe i autapomorfe nu poart informaie taxonomic. Statutul caracterului se poate modifica n dependen de nivelul ierarhic studiat: un caracter apomorf la un anumit nivel de investigare: un caracter apomorf la un anumit nivel de cercetare devine deja pleziomorf. De exemplu, membrele cu 5 degete ale omului, oprlelor i broatelor la compararea lor cu nottoarele petilor sunt un caracter sinapomorf i prezint o dovad elocvent a originii lor comune. Dar n acelai timp la unele animale terestre membrele posterioare au cte 4 degete (psrile), 2 degete (paricopitatele), sau 1 deget (imparicopitatele). n procesul de elucidare a legturilor filogenetice dintre oameni, amfibieni i cai de exeplu, n mod eronat se poate contrapune memrul cu 5 degete celui cu un deget.. dar dac este respectat principiul sinapomorfiei pericolul erorii dispare, deoarece apariia membrului uniddigitat este secundar apariiei membrului cu 5 degete, adic membrul cu un deget este apomorf fa de cel cu 5 degete. Din acest punct de vedere membrul cu 5 degete din punct de vedere al acestui nivel de cercetare este o sinpleziomorfie i nu trebuie luat n consideraie la construirea clasificrii. Este interesant relaia dintre principiul sinapomorfiei i noiunile de omologie i analogie. Noiunea de omologie exprim asemnarea determinat de originea comun. Se considr c prezena organelor omoloage este o dovad a apartenenei organismelor la unul i acelai taxon. Din exemplul relatat anterior reiese, c omoloage, adic determinate de originea comun pot fi att caracterele sinapomorfe, ct i cele sinpleziomorfe. Motiv pentru ntrunirea organismelor ntr-un taxon, ns, sunt doar sinapomorfiile. De aici reiese, c omologia n sine, nu poate servi drept baz pentru clasificare. Analogia exprim asemnarea, care nu este determinat de originea comun. n cladistic acest fenomen este numit homoplazie i se determin ca stare de apariie multipl a caracterului n cursul evoluiei. Principiul monofiliei Din punct de vedere al teoriei evoluioniste taxonul este considerat natural n cazul, n care el const din organisme cu originea comun. Deoarece viaa pe pmnt are origine unic, reiese, c
orice grut taxonomic este unul natural. Pentru a evita asemenea inconveniene Henning a fosrmulat principiul monofiliei stricte: Taxonul A este mai apropiat din punct de vedere al gradului de rudenie de taxonul B, comparativ cu taxonul C, dac A i B au un strmo comun, care nu este strmo i pentru C. n baza acestui principiu Henning deoasebea trei tipuri de grupuri taxonomice: strict monofiletice (holofiletice); parafiletice i polifiletice. n opinia lui Henning taxonul strict monofiletic include cel mai recent stmo i toi descendenii lui. Grupul monofiletic este determinat n baza sinapomorfiilor. Grupul parafiletic include strmoul comun i unii din descendenii lui. Un asemenea grup este determinat prin pleziomorfii. Grupurile polifiletice includ unii dintre descendenii strmoului comun , dar nu include strmoul comun. Aceste grupe se apreciaz n baza homoplaziilor. n cladistic sunt recunoscui doar taxonii monofiletici. Drept exemplu de grup parafiletic poate servi clasa Reptilelor, care care nu include psrile, adic nu include toi descendenii strmoului comun. Alt exemplu de taxon parafiletic ste filumul Chlorophyta (algele verzi), care nu include plantel superioare- descindenii direci ai acestui grup. Regnul Fungi este unul polifiletic, deoarece const din organisme, care nu provin de la un strmo comun. Exemplele de mai sus sunt inacceptabile din punct de vedere al cladisticii. Din punct de vedere al cladisticii Henning, grupurile monofiletice, parafiletice, polifiletice sunt determinate de atitudinea fa de originea comun. Mai trziu ali teoreticieni ai cladisticii, aa ca Platnick, 1979 i Patterson, 1982 au ajuns la concluzia, c din acest punct de vedere teoria cladisticii d bre n legtur cu faptul, c se bazeaz pe ceva, care nu este cunoscut originea comun. n opinia lor, formarea grupurilor taxonomice trebuie s aib loc n baza sinapomorfiilor, i abea dup aceasta, clasificaia obinut urmeaz a fi explicat din punct de vedere al filogeniei. Astfel, originea este exclus din analiz i devine doar parte component a discuiei tabloului relaiilor taxonomice (paternului) obinut. Aceast abordare a stat la baza aa numitei cladistici de patern. Principiul grupurilor nfrite Cu toate c deja am operat de mai multe ori cu noiunile de strmo i descendent, cladistica niciodat nu precaut taxonii care sunt supui clasificrii n calitate de strmoi ai unui oricare grup, chiar dac sunt cunoscute forme mai progresive nrudite cu el. n cladistic este recunoscut doar un criteriu de stabilire a grupurilor taxonomice sinapomorfia, care nu permite de a stabili relaia strm-urma.Deaceea din punct de vedere al cladisticii nu este corect de spus, c taxonul B provine de la taxonul A, ci c taxonii A i B au avut un strmo comun. Fa de care taxonul A cptat careva caractere apomorfe suplimentare, iat taxonul B nu. Taxonii ntre care exist asemenea relaii au fost numii de ctre Henning grupuri nfrite. El definea grupurile nfrite ca grupuri de specii, care provin de la strmoul grupului monofiletic n rezultatul unui proces de segregare, comun pentru acest grup. Grupurile nfrite sunt stabilite n baza sinapomorfiilor: Grupurile nfrite o pereche de taxoni, care au unul sau mai multe caractere unicale.
Principiul parsimoniei Una din particularitile cladisticii const n aplicarea principiului economiei sau parsimoniei. Acest principiu, numit deasemenea si briciul lui Occam a fost propus nc n sec.XIV de ctre filozoful englez WW.Occam. Principiul parsimoniei este parte component a metodei tiinificei se utilizeaz pentru excluderea presupunerilor, care sunt aplicate special pentru explicarea unui caz concret (adic a ipotezelor ad hoc). Asemenea ipoteze servesc pentru a putea ignora observaiile, care mrturisesc contra unei teorii concrete.Cu alte cuvinte, numrul de presupuneri, pe care urmeaz s le fac cercettorul pentru a explica observaiile, trebuie s fie minim.Ipotezele, care sunt utilizate extra acestui minim sunt ipoteze ad hoc.Aplicarea parsimoniei reiese din formularea iniial a metodelor filogenetice, dat de Henning, care considera c: Provenirea n rezultatul convergenei nu trebuie presupus a priori; adic dac lipsesc dovezile convergenei sau paralelismului, urmeaz a fi naintat ipoteza despre originea unic a acestui caracter la diferite organisme. Trebuie de menionat, c cea mai econom ipotez nu neaprat este cea mai aproape de realitate. Este vorba doar de o asemenea presupunere, la care trebuie s ne oprim, pentru a nu fi pui n condiiile, n care vom atrage ipoteze ad hoc. n corespundere cu principiul parsimoniei evoluia este recunoscut de ctre claditi ca ireversibil, unidirecionat i divirgent. Cazurile de evoluie n reea, simbiogeneza, radiaia adaptiv nu sunt analizate n acest context. Convergena (homeoplazia) este considerat drept eveniment puin probabil.Principiul evoluiei econome amintesc asemenea presupuneri ca gazul ideal sau corp absolut negru: sunt irealizabile n realitate, dar foarte utile n cercetare. Etapele analizei cladistice Analiza cladistic bazat pe principiile expuse mai sus include urmtoarele etape: 1. La prima etap se efectuiaz codificarea caracterelor , care se includ n matrice. O particularitate a metodei cladistice const n aceea, c pe primul loc sunt plasate caracterele calitative. Indicatorii metrici n cadrul analizei cladistice sunt plasai n grupuri , ca de exemplu: mic, mediu, mare. Codificarea adecvat a caracterelor este o problem separat foarte serioas. De faptul, ct de exact codificarea reflect strile existente ale caracterului, depinde ct de reale vor fi rezultatele analizei. Vom examina cteva tipuri de caractere i metode de codificare. Presupunem c trebuie s descriem culoareacorolei la dou specii de plante. Fie c la specia A corola este de culoare roie, iar la specia B alb. Aceast informaie poate fi codificat n form binar: starea caracterului la specia A este codificat cu 0, iar la specia B cu 1. Caracterele care au doar dou stri posibile se numesc caractere binare. La codarea binar nu are importan cum se noteaz fiecare dintre strile posibile ale caracterului cu 0 sau cu 1. Valorile pot fi schimbate, fr a schimba sensul codificrii. Tabel. Codificarea binar a caracterului culoarea corolei la doi taxoni taxonul Caracterul Codul A Corol roie 0 B Corol alb 1
Atunci, cnd caracterul are multiple forme (nu numai dou) se aplic un alt tip de codificare codificarea aditiv. Tabel. Codificarea aditiv a caracterului culoarea corolei la trei taxoni taxonul Caracterul Codul A Corol roie 0 B Corol albastr 1 C Corola albastr 2 La codarea unui caractre este necesar de a stabili dac acesta este ordonat sau neordonat. n cazul caracterelor ordonate, caracterul formeaz succesiuni de tipul alb-gri-negru sau sfericelipsoidal-cilindric. n cazul de mai sus nu exist un ir ordonat. Caracterele ordonate se codeaz n mod deosebit. De exemplu caracterul, irul ordonat al strilor cruia este 0-1-2, unde 1 este starea intermediar a caracterului, poate fi codificat astfel: 00-01-11. O asemenea codificare poart numele de codificare aditiv. Tabel. Codificarea aditiv a caracterului ordonat taxonul Caracterul Codul A Frunz rotungit 00 B Frunz ovoid 01 C Frunz n form de inim 11 Transformarea n irul caracterului ordonat poate fi liniar (ca n exemplele de mai sus) sau ramificat. Ultimile deasemenea pot fi codificate n sistema aditiv binar C A B D Tabelul. Codarea aditiv binar a transformrii ntre 4 stri ale caracterului Taxonii Codul A 000 B 001 C 011 d 101 (de exemplu: A- 000- sistemul circulator la petii osoi cu 4 perechi de artere branhiale; B001- sistem circulator la reptile cu 2 arcuri ale aortei; C-011 sistemul circulator la mamifere cu reducerea arcului drept al aortei; D-101 sistemul circulator la psri cu reducerea arcului stng al aortei) n cazul caracterelor binare transformarea caracterului poate avea loc n ambele direcii. n cazul n care direcia transformrii este srabilit, atunci caracterul este polarizat. Polarizarea nu trebuie s fie confundat cu irul caracterelor orientate. De exemplu irul 0-1-2 poate fi polarizat n trei moduri diferite De la 0 la 1 la 2 De la 2 la 1 la0 De la 1 la 2 i de la 1 la 0. n cazul caracterelor polarizate n analiz se ia doar varianta care reflect polaritatea
2. Dup ce a fost alctuit matricea, programa ncepe selectarea cilor potenial posibile de transformarea evoluionist a fenotipului ori genotipului iniial n toate celelalte pe calea modificrii pe rnd a strilor tuturor caracterelor. n rezultat se construiesc cladograme (sau arbori), care constau din linii, numite ramuri. Locurile de intersectare a ramurilor se numesc noduri. Vrfurile ramurilor corespund taxonilor, care au fost inclui n analiz. Arborii pot fi nenrdcinai (numii reele ori dendrite) i nrdcinai (anume acetea sunt cladograme). Arborii nenrdcinai reflect natura legturilor dintre taxoni, dar nu pot da informaie despre transformarea caracterelor.
Fig. Arbore nenrdcinat (dendrit) a); arbore nrdcinat (cladogram) b) Pentru a transforma dendrita n arbore nrdcinat se efectuiaz nrdcinarea (rooting) ei. nrdcinarea poate fi direct i indirect. nrdcinarea indirect se face prin ntroducerea n analiz a unui grup expert (outgroup) un taxon, care din start este separat de totalitatea taxonilor analizai (ingroup). Este de dorit, ca grupa extern s nu posede apomorfii caracteristice pentru ingrup. n rezultatul nrdcinrii transformarea a cel puin un caracter capt polaritate. Uneori se polarizeaz mai muli factori, ori chiar toi. Aa dar, polaritatea caracterului se citete numai depe arbori nrdcinai. Metoda direct de determinare a polaritii necesit utilizarea datelor ontogenetice: n cazul transformrii ontogenetice a caracterului de la starea mai rspdit la cea mai puin rspndit, starea mai rspndit se apreciaz ca pleziomorf, iar cea mai puin rspndit apomorf. De exemplu fantele branhiale se ntlnesc la embrionii tuturor vertebratelor i la indivizii maturi ai petilor, dar nu la indivizii maturi ai tetrapozilor. De aici reiese, c starea
mai rspndit (prezena fantelor) prezint o pleziomorfie, iar transformarea fantelor n alte structuri prezint apomorfii. 3. Dintre multitudinea de cladograme este necesar de a le selecta pe cele mai econome (care corespund principiului parsimoniei). Deoarece numrul de cladograme generate este foarte mare, pentru efectuarea seleciei se utilizeaz cteva metode. Cutarea exhaustiv (se precaut toate cladogramele posibile). Se aplic doar pentru grupuri mici i un numr mic de caractere. Metoda ramificaiei i limitrii (branch-and-bound method, BAB). Se construete cladograma etalon. Lungimea ei este luat n lucru ca maximal admisibil. Cladogramele cu lungime mai mare se exclud. Cutarea euristic. Cercettorul fixeaz un numr minimal de cladograme, care urmeaz a fi generate. De obicei nu mai puin de 10000, dup care se lucreaz doar cu aceti arbori. Cutarea euristic se efectuiaz n dou etape: 1.adugarea pas cu pas, care determin consecutivitatea adugrii taxonilor la arborele ce se construiete, 2. schimbul de ramuri diferite variante de schimb cu locul a ramuriilor arborelui obinut la prima etap. Adugarea pas cu pas poate fi efectuat prin mai multe metode: Se aleg 3 taxoni si se alctuiete reeaua de baz. Taxonii rmai se adaug pas cu pas la fiecare din ramurele arborelui in construcie si se calculeaz distana pn la el. Se selecteaz varianta de adugare cu cea mai mic lungime. Algoritmul simplu (Farris). Se ia un taxon, de obicei primul din list, dup care se calculeaz gradul de diferen dintre el si toi ceilali taxoni sub form de sum a deosebirilor absolute ntre caracterele lor. Acest indice se numete indice de avansare. Se construete se construete structura de baz ce const din taxonul de baz i ali 2 cei mai apropiai (cu cel mai mic indice de avansare), apoi se adaug ceilali taxoni n ordinea creterii indicelui. Procedura cea mai amnunit elaborat de Swofford presupune aplicarea unei proceduri dinamice, care const n aceea, c succesiunea adugrii taxonilor se revede pe parcursul construirii arborelui. n rezultatul adugrii pas cu pas se obin ameliorri eseniale de ordin local. Dentru a mbunti esenial calitatea cladogramei se aplic schibrul ntre elemente perfectate ale arborelui obinut adic cea de-a doua etap schimbul de ramuri. Schimbul de ramuri poate fi realizat prin mai multe metode: Schimbul ntre vecinii apropiai (nearest neighbour interchange) sau schimb local de ramuri;
Fig. Exemplu de schimb de ramuri prin metoda schimbului ntre vecinii apropiai (schimbul local). Ramura luat n cercetare (desenul de sus)are la ambele extremiti cte o pereche de vecini (A+B; C; D; E+F). Unul din ei C este schimbat cu fiecare dintre vecinii de pe partea opus, ceea ce genereaz noi cladograme, Retezare i altoire (subtree pruning and regrafting); de la cladograma obinut se separ o subcladogram nrdcinat. Aceasta apoi se alipete la fiecare dintre ramurile claogramei rmase. Se apreciaz lungimea topologiei obinute i se selecteaz cele cu lungime minim. Despicare i reunire (tree bisection and reconnection). Subladograma secionat se transform n reea, iar dup asta din nou se nrdcineaz pe fiecare dintre celelalte ramuri rmase. A B
Fig. Exemple de Retezare i altoire A i Despicare i reunire B.
4. Din multitudinea de cladograme se selecteaz cele mai laconice, care includ cel mai mic numr de pai evolutivi, adic modificri ale strii caracterului. La aceast etap are loc cntrirea ipotezelor filogenetice. La aceast etap o importan major o are tipul de interaciuni dintre caracterele taxonilor. Acestea pot fi: Cogruente se comport la fel; Compatibile nu sunt contradictorii (determin grupuri diferite din aceiai categorie) Contradictorii Matrice i cladogram cu caractere congruente
Matrice i cladogram cu caractere contradictorii.
Atunci cnd se evideniaz elemente contradictorii este necesar de a efectua optimizarea. Exist mai multe metode care permit de a efectua optimizarea: Optimizarea aditiv (analiza caracterelor nepolarizate ordonate) Optimizare neaditiv (analiza caracterelor neordonate, nepolarizate) Optimizarea Dollo (caractere ordonate polarizate sau nu) Optimizarea Camin-Sokal (cactere polarizate ordonate) 5. Construirea cladogramei de consens din cteva cladograme obinute n rezultatul optimizrii. Exist 3 procedee de baz de construire a cladogramei de consens:
Consensul strict n arborele de consens se includ doar acele sectoare, care sunt prezente n toate cladogramele elaborate, iar taxonii, legturile dintre care sunt contradictorii se exprim prin linii paralele. Consensul majoritii n arborele final se includ segmentele, care sunt cuprinse n mai mult de 50% din cladogramele generate la etapa precedent.
Cladograme obinute n baza a trei cele mai scurte cladograme n conformitate cu principiul consensului strict i consensului majoritii. Consensul dup Adams. Componentele nestabile sunt transferate n nodul cel mai apropiat care le unete.
6. Cladograma de consens se apreciaz din punct de vedere al stabilitii. Pentru aceasta se aplic 2 metode: bootstrap i jackknife.
Metoda Bootstrap se bazeaz pe alctuirea pseudomatricelor de aceleai dimensiuni Metoda Jackknife const n expluderea unor date din matrice.Cladograma de consens este verificat din punct de vedere al gradului de corspundere. n cladistic prin acest termen se exprim gradul de corespundere a ntregului complex de date examinate cladogramei obinute.
Fig. Etapele metodei Bootstrap: a este construit cea mai econom cladogram pentru un ansamblu iniial de factori. - se genereaz o pseudomatrice de aceiai dimensiune, din care pot lipsi anumite date, iar altele se pot repeta. se construiesc cladograme n baza pseudomatricelor i la final se obine unarbore de consens dup principiul consensului majoritii. Procentul cladogramelor parsimonice , n care se ntlnete aceast grup se indic la baza ei si reflect credibilitatea lor. 7. Aprecierea corespunderii cladogramei complexului de caractere analizate. n calitate demsur a corespunderii pentru datele discrete este aplicat indicele de consisten (ci consistency index), care se calculeaz dup formula:
Unde s este numrul real de modificri ale strii caracterului, iar m- este numrul minim de modificri ale strii caracterului, Ci variaz ntre 1 pentru corespunderea absolut i 0 pentru lipsa corespunderii. Indicele consistenei poate fi aplicat att pentru un caracter, ct i pentru ansamblul de caractere n cladogram. n ultimul caz vorbim despre Indicele Total de Consisten
n calitate de criteriu de corespundere a cladogramei obinute polarizrii tuturor caracterelor de la starea pleziomorf la cea apomorf este utilizat indicele de retenie (ri - retention index). Pentru un caracter particular acest indice se calculeaz conform formulei:
In care g- numrul maximal de pai posibili pentru caracterul dat, s numrul real de pai, m/ numrul minimal de pai posibili. Indicele total de retenie , care caracterizeaz toate caracterele cladogramei se calculeaz dup formula
Dac ri=1 aceasta arat c polaritatea caracterelor este reflectat totalmente pe cladogram, iar valorile mai mici de unitate indic asupra posibilitii de dispariie secundar a caracterelor apomorfe.. 8. Dup ce a fost efectuat evaluarea cladogramei de consens dup indicii de mai sus, nodurilor acestei cladograme le este atribuit un anumit rang taxonomic. n aa mod are loc formarea unei clasificri ierarhice a organismelor cercetate.
Avantajele metodei cladistice: A ntrodus abordri noi n sistematic, determinnd de fapt aspectul contemporan al,acestei tiine. A introdus termenii de cladogram, apomorfie, iar interdicia pentru utilizarea taxonilor poli- i parafiletici pentru construirea cladogramelor i-a gsit aplicare i dincolo de limitele metodei cladistice, devenind un principiu general pentru taxonomia contemporan. Anume cladistica a creat baza metodologic pentru analiza genotipic, fiind pn nu demult unica metod de analiz filogenetic a secvenelor nucleotidice. Dezavantajele metodei cladistice: Aprecierea ipotezelor n conformitate cu principiul parsimoniei nu este o garanie contra erorilor, deoarece excluderea general a unei ipoteze n baza faptului c aceasta este puin posibil nu este chiar corect. Metodele de construire a cladogramei de consens nu face fa atunci cnd poziia taxonilor este echivalent. Refuzul de a recunoate taxonii contemporani n calitate de taxoni strmoi nu este raional ntotdeauna deoarece sunt cazuri, cnd ntre doi taxini contemporani sunt stabilite relaii sigure de tipul strmo - descendent. Analiza cladistic n baza fenotipului nu este n stare de a nltura toate problemele legate de fenomenul de convergen. n cadrul taxonilor, n care formarea convergent repetat a caracterelor, este o regul, ca de exemplu, la protiste, cladistica simplific neargumentat tabloul modificrilor evolutive. Fenomenele de evoluie n reea, radiaiei
adaptive multiple i n general procesul de microevoluie n populaie nu pot fi descrise n mod adecvat utiliznd posibilitile cladisticii.
METODE STATISTICE N ANALIZA FILOGENETIC
Unul din postulatele principale ale cladisticii, principiul parsimoniei, este n acelai timp i unul din cele mai controversate postulate. Metoda parsimoniei iniial elaborat pentru caracterele morfologice, prezint mai multe versiuni (Wiley, 1981; Felsenstein, 1982; Wiley et al., 1991; Maddison i Maddison, 1992; Swofford i Begle 1993). Eck i Dayhoff n 1996 par a fi fost primii care au utilizat aceast metod pentru realizarea arborilor, pornind de la secvene de aminoacizi. Fitch (1971) i Hartigan (1973) au elaborat un algoritm mai riguros al metodei MP pentru secvene de nucleotide. n cadrul acestei metode se iau n considerare 4 sau mai multe secvene aliniate de nucleotide sau aminoacizi (m4), iar nucleotidele (aminoacizii) taxonilor ancestrali sunt luate n considerare separat pentru fiecare site i o anumit topologie, presupunnd c modificrile produse de mutaii apar la nivelul oricreia dintre cele patru nucleotide (sau 20 de aminoacizi). Se calculeaz apoi, cel mai mic numr de substituii nucleotidice (sau de aminoacizi) care explic ntregul proces evolutiv pentru o anumit topologie. Acest calcul este realizat pentru toate topologiile presupuse a fi corecte i topologia care implic cel mai mic numr de substituii este aleas ca reprezentnd arborele corect. Cu ct este implicat mai puin procesul evolutiv de creare a unei inferene filogenetice, cu att putem obine concluzii mai exacte. Dac nu exist substituii n paralel sau n sens invers (nu exist omologie) la fiecare site nucleotidic i numrul de nucleotide examinate (n) este foarte mare, ar trebui ca utiliznd MP s se obin arborele real. Cu toate acestea, n practic, secvenele de nucleotide sufer adesea substituii n paralel sau n sens invers, i n este relativ mic. n acest caz, MP tinde s confere o topologie incorect. n plus, Felsenstein (1981) a artat c atunci cnd rata substituiei nucleotidelor variaz proporional cu direcia evolutiv, MP poate genera topologii incorecte chiar i atunci cnd se examineaz un numr foarte mare de nucleotide. n anumite condiii, aceasta se poate ntmpla chiar i atunci cnd rata de substituie este constant pentru toate direciile evolutive (Hendy i Penny, 1989; Takezaki i Nei, 1996). n acest caz, ramurile lungi (sau cele scurte) ale arborelui real, tind s se alture, sau se atrag n cadrul arborelui final. Astfel, acest fenomen, este adesea numit long branch attraction (engl. atracia dintre ramurile lungi) (Hendy i Penny, 1989) sau Short branch attraction (engl. atracia dintre braele scurte) (Nei, 1996). n analiza parsimoniei este de asemenea dificil abordarea inferenei filogenetice ntr-un cadru statistic, deoarece nu exist o cale de calcul a mediei i varianei numrului minim de substituii obinute pe criteriul parsimoniei. Astfel, utilizarea metodei parsimoniei este realist nu din cauza, c ar da o impresie real despre mersul evoluiei, ci pentru c permite de a evita presupunerile, care nc nu au confirmare. Parsimonia este un procedeu de evaluare a puterii de explicare a ipotezelor, i nu un procedeu de apreciere realitii ei.
Aceast situaie impune necesitatea de a combina arsenalul analitic al cladisticii cu mijloacele statisticii matematice. Astfel au aprut metodele statistice ale analizei filogenetice, care n prezent sunt ntruchipate n dou variaii : verosimilitudine maxim i analiza Bayes.
Metoda verosimilitudinii maxime (maximum likelihood) Analizele filogenetice tind s deduc istoria evolutiv (sau un set de istorii probabile) care corespunde cel mai bine setului de date observat (n cazul de fat este vorba de secvene nucleotidice sau de aminoacizi dar poate fi vorba i de caractere morfologice, frecvene ale genelor, situsuri de restricie etc.). Necunoscutele problemei sunt ordinea de ramificare i lungimea ramurilor a filogeniei. Pentru a aplica metodele ML este nevoie de un model concret de substituie ce descrie transformarea unei secvene n alta. Metodele ML de reconstrucie filogenetic evalueaz ipoteza despre istoria evolutiv n termeni probabilistici (Care este probabilitatea c o anumit istorie evolutiv - topologie i un anumit model de substituie vor da natere datelor observate). O istorie evolutiv ce are o probabilitate mai mare de a da natere datelor observate are prioritate fa de una cu o probabilitate mai mic. Aceast metod a fost utilizat pentru prima dat n reconstruciile filogenetice de ctre Cavalli-Sforza i Edwards n 1967 ns aceti autori au considerat calculele necesare prea complicate pentru calculatoarele de la acea vreme i au dezvoltat metode aproximative ca de exemplu ME (Huelsenbeck & Crandall, 1997). Felsenstein a utilizat pentru prima dat aceast metod n 1981 pentru analiza filogenetic a secvenelor de nucleotide, dup care metoda a nceput s fie din ce n ce mai utilizat. Printre avantajele acestei metode se numr variana mic i posibilitatea utilizrii unui numr minim de parametri. Chiar pentru un numr mic de nucleotide acest tip de metode depesc de multe ori metodele bazate pe distane i cele MP. Principiul de baz al metodei implic calcularea verosimilitudinii unei filogenii. Deoarece majoritatea modelelor de substituie folosite sunt reversibile n timp, verosimilitudinea unei filogenii este independent de localizarea rdcinii. Presupunnd c fiecare situs evolueaz independent putem calcula verosimilitudinea fiecrui situs separat i s combinm aceste valori pentru obinerea unei valori finale. Pentru a calcula verosimilitudinea unui situs j trebuie s lum n considerare toate scenariile posibile prin care starea final a situsului ar fi putut evolua. Bineneles c unele scenarii sunt mult mai plauzibile dect altele, ns fiecare are o anumit ans s se fi produs. Spre exemplu dac considerm o filogenie n rdcin am fi putut avea teoretic oricare din cele 4 nucleotide (A, G, C, T), pentru oricare alt nod intern este de asemeni posibil existena oricrui nucleotid. Deci avem un total de 4 X 4 = 16 posibiliti. Cum oricare dintre aceste 16 scenarii ar fi putut duce la setul final de date observate, probabilitile tuturor evenimentelor trebuiesc calculate i nsumate pentru a obine probabilitatea unui situs j (aceast probabilitate depinde dup cum am mai menionat de topologie i de modelul de substituie nucleotidic). Cum am presupus c fiecare situs evolueaz independent de celelalte, probabilitatea unui scenariu evolutiv este egal cu produsul probabilitilor tuturor situsurilor 1, 2, 3, j n. Pentru c probabilitatea unei singure observaii este foarte mic n locul lor se utilizeaz logaritmii acestora, deci probabilitile sunt acumulate ca sum a logaritmilor verosimilitudinilor fiecrui situs luat n parte.
Din cauza volumului foarte mare de calcule ce trebuiesc efectuate dac numrul de taxoni este mai mare ca 10 i secvenele utilizate sunt mari au fost dezvoltate o serie de algoritmi de cutare (ca i n cazul metodelor MP). Cum modalitatea real de substituie a nucleotidelor este foarte complicat, s-ar putea crede c un model matematic cu muli parametri este mai bun dect un model cu mai puini parametri. Totui chiar dac un model cu mai muli parametric corespunde datelor mai exact dect unul mai simplu, numrul mare de parametri l face mai sensibil la erori. Deci este mai util utilizarea unui model mai simplu atta timp ct acesta aproximeaz modul de substituie al nucleotidelor suficient de bine. n cazul metodelor ML corespondena dintre model i date poate fi examinat folosind testul LR (Likelihood Ratio Test) sau criteriul informaiei al lui Akaike (AIC Akaike Information Criterion). Cnd avem dou modele 1 i 2 i modelul 1 este caz particular al modelului 2, iar topologia corect este cunoscut putem calcula testul LR dup formula LR=2(lnL2-lnL1) unde lnL1 i lnL2 sunt valorile ML pentru modelele 1 i 2. n acest mod putem testa dac modelul 2 este semnificativ mai bun dect modelul 1.
ASPECTE ALE EVOLUIEI MOLECULARE EVOLUIA SECVENELOR NUCLEOTIDICE I PROTEICE De la Charles Darwin ncoace reconstruirea istoriei evolutive a vieii i redarea acesteia sub forma unui arbore filogenetic a fost un vis pentru muli biologi , Haeckel (1866) fiind primul care a ncercat sa realizeze acest lucru. Modul ideal pentru realizarea acestei sarcini imense este utilizarea registrului fosil, dar ntruct acesta este fragmentar muli cercettori au ncercat s foloseasc morfologia i fiziologia comparat. Folosind aceste dou metode s-a reuit deducerea multor aspecte importante din istoria vieii. ns schimbrile morfologice i fiziologice fiind extrem de complexe, iar aprecierea importanei evolutive a unui caracter fiind de cele mai multe ori subiective, arborii filogenetici construii au fost ntotdeauna subiect pentru controverse. n ultimul timp datorit realizrilor biologiei moleculare situaia s-a schimbat drastic. Dac n anul 1982 n GenBank erau doar 606 secvene (680.338 pb) aceast baz de date fiind la nceputurile sale, iar n 1997 se ajunsese dup 15 ani abia la 1.765.847 de secvene (1,160,300,687pb) astzi baza de date numr 46 947 388 de secvene cu un total de aproximativ 51 674 486 881 pb de la peste 165 000 de specii (Benson, 2005). Aceast explozie se datoreaz n primul rnd perfecionrii metodelor de secveniere i a dus la apariia unui mare numr de studii de evoluie molecular. Planul structurii tuturor organismelor fiind nscris n ADN (sau ARN n cazul unor virusuri) apare posibilitatea de a studia relaiile evolutive dintre organisme comparndu-le ADN-ul. Acest mod de a privi lucrurile are cteva avantaje fa de cele clasice ce utilizeaz caractere morfologice i fiziologice: (1) Macromolecula de ADN este format din patru tipuri de nucleotide: adenina (A), timina (T), citozina (C) i guanina (G) n ntreaga lume vie putnd fi folosit astfel pentru a compara orice grupe
de organisme, plante, bacterii, arhebacterii, animale, fungi. n metodele tradiionale aceasta este pur i simplu imposibil. (2) ntruct modificrile evolutive ale ADN urmeaz un model mai mult sau mai puin regulat este posibil utilizarea modelelor matematice pentru a formula modificrile i compara ADN-ul de la organisme foarte ndeprtate din punct de vedere filogenetic. Schimbrile morfologice sau fiziologice urmeaz modele extrem de complexe chiar dac analizm perioade scurte de timp nefiind clar dac diferitele aproximri necesare pentru realizarea unor filogenii sunt reale sau nu. (3) Genomurile diferitelor organisme constau din lungi secvene de nucleotide (de exemplu genomul uman are 9x1012 pb) coninnd astfel cantiti mult mai mari de caractere cu importan filogenetic spre deosebire de alte tipuri de date. Taxonomia este una dintre cele mai controversate ramuri din biologie, definiiile speciilor, genurilor familiilor etc. fiind adesea subiective, dezacordul dintre specialiti privitor la statutul unui anumit taxon fiind ceva obinuit. Filogeneza este mai puin controversat dect taxonomia n general, ea dorind n primul rnd s rezolve problema relaiilor dintre organisme, iar statutul sistematic al organismelor are o importan secundar. Cele dou domenii sunt ns strict corelate, clasificarea organismelor trebuind s reflecte istoria lor evolutiv (Ax, 1987). n acest sens filogenetica molecular joac un rol important n dezvoltarea bazelor tiinifice ale taxonomiei, ns nu trebuie privit ca atotputernic existnd cazuri cnd unele probleme din nomenclatur nu pot fi rezolvate pe aceast cale. Un exemplu interesant de utilizarea a datelor moleculare n stabilirea relaiilor filogenetice dintre diveri taxoni l constituie cetaceele a cror legturi cu celelalte ordine de mamifere au fost mult discutate. Prin analiza evoluiei genei fibrinogenului ct i prin datele referitoare la inseria n genom a SINE (short interspersed repetitive elements) ce pot fi privite ca caractere sinapomorfe (Nei & Kumar, 2000) s-a ajuns la concluzia c printre mamiferele actuale, cele mai nrudite cu cetaceele sunt hipopotamii. Pe lng faptul c la acealeai rezultate s-a ajuns prin dou metode absolut diferite, ipoteza este susinut i de datele paleontologice care indic nrudirea cetaceelor cu mezonichidele, un grup de ungulate carnivore din oligocen (Benton,1997). METODE DE DEDUCERE A DIRECIILOR DE FILIAIE PORNIND DE LA DATE MOLECULARE Deducerea informaiilor filogenetice din datele moleculare necesit alegerea metodei potrivite din multele disponibile. Construirea unei ipoteze filogenetice este un procedeu de estimare, adic se face o estimare optim a unei filogenii bazndu-ne pe informaia incomplet coninut n datele disponibile. n contextul sistematicii moleculare de obicei nu avem informaii directe despre trecut avem acces doar la organisme i molecule actuale. Deoarece putem gsi un scenariu evolutiv prin care orice filogenie poate fi explicat e nevoie de un criteriu pentru a selecta una sau cteva filogenii preferate dintr-un set de filogenii posibile. Metodele de deducere filogenetic ncearc s realizeze acest scop n unul din dou moduri: (1) definind o secven specific de pai (un algoritm) ce duce la obinerea unui arbore filogenetic; (2) definind un criteriu pentru a compara filogeniile alternative ntre ele i deciznd care e mai bun.
Metodele pur algoritmice combin deducerea arborelui filogenetic i definiia arborelui preferat ntr-o singur supoziie. Aceste metode includ toate formele de analiz a grupelor perechi (cum ar fi UPMGA) i alte metode bazate pe distane cum ar fi NJ (neighbor joining). Ele sunt de obicei mai rapide deoarece merg direct spre arborele final fr a necesita evaluarea unui numr mare de soluii. A doua grup de metode presupune doi pai logici. Primul este definirea unui criteriu de optimalitate (descris formal de printr-o funcie obiectiv) cu ajutorul cruia fiecrui arborie i se acord un punctaj folosit pentru compararea arborilor unul cu cellalt. Al doilea pas este utilizarea unui algoritm specific pentru calcularea valorii funciei obiective i gsirea arborelui cu cel mai bun punctaj. Astfel supoziiile evolutive fcute n primul pas sunt decuplate de folosirea calculatorului n pasul al doilea. Preul acestei clariti logice este viteza lent de lucru a calculatorului, iar pentru seturi date ce conin mai mult de 20 de taxoni arborele optim nu poate fi ntotdeauna gsit din cauza numrului mare de posibile soluii. Folosirea algoritmilor este radical diferit ntre cele dou metode. n cazul metodelor pur algoritmice, algoritmul este cel care definete criteriul de selecie al arborilor i are deci o valoare crucial. n metodele bazate pe un criteriu algoritmul este doar unealt pentru evaluarea valorii funciei obiective i cutarea arborilor ce optimizeaz aceast valoare. Deoarece metodele bazate pe criterii acord fiecrui arbore analizat un punctaj, filogeniile pot fi aranjate n ordinea preferinelor. Aceasta reprezint un avantaj enorm n faa metodelor pur algoritmice. Dac un set de date poate fi explicat egal de bine de cteva mii de arbori filogenetici, folosind metode bazate pe criterii nu vom fi dui n eroare c un anumit arbore explic datele foarte bine. O metod algoritmic d un singur arbore, neputnd ti ct confiden putem avea n acesta. Unii autori (Hedges et al., 1992) au sugerat utilizarea metodelor statistice cum ar fi bootstrapping-ul pentru determinarea gradului de confiden pe care l putem avea ntr-un arbore gsit prin metode pur algoritmice. EVOLUIA SECVENELOR DE AMINOACIZI nainte de inventarea metodelor rapide de secveniere n 1977 (Maxam & Gilbert, 1977; Sanger et al., 1977) majoritatea studiilor de evoluie molecular se bazau pe secvene proteice. Astzi secvenierea ADN-ului este mult mai simpl i rapid dect secvenierea proteinelor i majoritatea secvenelor proteice sunt obinute prin transcriere folosind codul genetic. Totui secvenele proteice i-au pstrat utilitatea pentru studiile filogenetice : ele sunt mult mai conservate, dect secvenele de ADN i pot oferi informaii utile n cazul comparrii grupelor ndeprtate din punct de vedere filogenetic. n plus, modelele matematice ale schimbrii evolutive a aminoacizilor sunt mult mai simple dect cele pentru secvene de acizi nucleici, motiv pentru care facem mai nti o prezentare a evoluiei secvenelor proteice i doar apoi a celor nucleotidice. Studiul schimbrilor evolutive al secvenelor polipeptidice ncepe prin compararea a dou sau mai multe secvene. Cel mai simplu indice este numrul de aminoacizi diferii (nd). Daca secvenele aliniate conin inserii sau deleii acestea trebuiesc eliminate nainte de calcularea nd. Un alt indice simplu este distana p ce reprezint proporia de aminoacizi prin care dou secvene
de aminoacizi difer. Se calculeaz mprind numrul de aminoacizi diferii la numrul total de situsuri comparate. p=nd/n Acest indice nu face nici o corecie pentru substituii multiple n cadrul aceluiai situs sau pentru diferenele n ratele de evoluie ntre situsuri. Urmtorul indice este distana cu corecie Poisson (PC) deoarece n realitate n cadrul aceluiai situs pot avea loc substituii multiple ceea ce duce la o relaie neliniar ntre p i timpul t. Odat cu trecerea timpului discrepana dintre nd i numrul real de substituii crete. Dac r este rata de substituie per an i per situs (pentru simplitate se presupune c e acelai pentru toate situsurile) numrul mediu de substituii per situs ntr-o perioada de timp t este rt, iar probabilitatea a k substituii (k = 1, 2, 3, ) este dat de distribuia Poisson : PC(k;t) = e-rt(rt)k/k! Pentru calcularea ratei de substituie a aminoacizilor folosim formula r= d/2t, unde d numrul de substituii per situs. Pentru a putea aplica formula trebuie s cunoatem momentul nceperii divergenei dintre cele dou secvene. Daca cunoatem rata de substituie a aminoacizilor putem folosi formula de mai sus pentru a estima timpul evolutiv t= d / (2r). n modul de calculare a distanei descris mai sus se presupune c rata de substituie aminoacizilor este identic pentru toate situsurile. Aceast aproximare de cele mai multe ori nu este valabil, rata substituiei fiind mai ridicat pentru situsurile mai puin importante din punct de vedere funcional, iar aminoacizii din centrul activ al unei proteine pot fi conservai pentru perioade de timp foarte mari (Kimura, 1983). Uzzel & Corbin (1971) au artat c de fapt distribuia numrului de substituii per situs k are o varian mai mare dect cea a distribuiei Poisson i urmrete de fapt o distribuie negativ binomial, iar distribuia lui r per situs urmrete o distribuie gamma. Forma distribuiei gamma -f(r) este dat de a care se mai numete i parametru de form sau parametru gamma. Distribuia gamma este foarte flexibil i ia diverse forme n dependen de a. Pentru a =, r este acelai pentru toate situsurile, pentru a=1, r urmeaz o distribuie exponenial indicnd c rata de substituie variaz mult de la un situs la altul, iar pentru a < 1, r are o distribuie i mai concav indicnd c un mare numr de situsuri au r = 0 MUTAIILE I RATA DE SUBSTITUIE A AMINOACIZILOR
Genele sau secvene ale moleculelor de ADN care funcioneaz ca modele structurale pentru ARNm sunt numite gene structurale. De cnd secvenele de aminoacizi din structura unui lan polipeptidic se pot determina cu ajutorul secvenelor nucleotidice din genele structurale, orice schimbare n secvena de aminoacizi este cauzat de mutaiile produse la nivelul ADN. De asemenea, o mutaie care produce o modificare n structura ADN nu este necesar s se reflecte n modificarea secvenei aminoacizilor, fapt datorat degenerrii codului genetic. naintea inventrii metodelor rapide de secveniere a ADN n 1977 (Maxam i Gilbert, 1977; Sanger et al., 1977), cea mai mare parte a studiilor de evoluie molecular au avut la baz utilizarea secvenelor de aminoacizi. Unele principii importante ale evoluiei moleculare, cum ar fi: evoluia prin duplicarea genelor (Ingram 1963; Ohno, 1970) i ceasul evoluiei moleculare (Zuckerkandl i Pauling, 1962; Margoliash, 1963), au fost descoperite prin studiul secvenelor de aminoacizi. n prezent, secvenierea ADN este mult mai simpl dect secvenierea aminoacizilor care este de obicei dedus din secvenierea nucleotidelor prin utilizarea codului genetic (Nei i Kumar, 2000). Nu este adevrat faptul c orice mutaie poate fi ncorporat ntr-o secven, deoarece speciile sunt alctuite din populaii cu numeroi indivizi, iar o mutaie nou aprut ntr-un individ, poate dispare din populaie prin ans sau prin aciunea seleciei purificatoare. Numai n cazul n care mutaia survine la nivelul ntregii populaii, atunci aceasta poate fi inclus n genomul speciei. Acest proces poart numele de fixare a mutaiei n populaie. Odat fixat o mutaie n cadrul unei populaii, orice individ aparinnd acelei populaii va avea aceeai mutaie. Cnd sunt comparate dou secvene de aminoacizi provenite de la dou specii diferite, sunt urmrii iniial, noii aminoacizi care au fost ncorporai. Cnd apare o mutaie n cadrul populaiei, supravieuirea alelelor depinde n principal de ans, dac este sau nu avantajoas din punct de vedere selectiv i de mrimea populaiei. De exemplu, considernd A1 gena slbatic i A2 alela, ntr-un organism diploid, gena mutant apare iniial n stare heterozigot (A1A2). n urma ncrucirii A1A2 x A1A1, dac nu rezult descendeni (generaia F1) din diferite cauze biologice (de exemplu sterilitatea homozigotului A1A1), gena mutant va dispare n generaia urmtoare. Supravieuirea genei alele, nu este sigur nici chiar dac n urma ncrucirii A1A2 x A1A1 se produc urmai, deoarece descendenii cu genotipul A1A2 apar cu o probabilitate de 0,5. n plus, exist i o ans de 0,25 ca descendenii cu aceste genom s nu apar (Nei i Kumar, 2000). Cnd o mutaie este neutr sau nu afecteaz fitness-ul individului purttor, frecvena relativ a mutantei poate crete sau descrete prin ans n cadrul populaiei (Nei i Kumar, 2000). Pn acum n toate modelele descrise s-a vorbit despre substituiile aminoacizilor de parc fiecare substituie este fixat n secvena considerat. De fapt secvenele de aminoacizi nu sunt independente, fiecare specie constnd din populaii, iar mutaiile au loc n individ ele putnd dispare prin deriv genetic sau prin selecie stabilizatoare. Doar cnd mutaia se rspndete n ntreaga populaie putem spune c ea este fixat. Cnd comparm dou secvene de aminoacizi de la dou specii observm doar schimbrile ce au fost fixate n genofondul speciei. Cnd o mutaie este neutr i nu afecteaz fitnesul indivizilor frecvena relativ (x) a alelei mutante poate crete sau descrete la ntmplare n interiorul populaiei. La un organism cu generaii discrete (cum ar fi insectele sau plantele anuale) i numr N de indivizi aduli frecvena unei alele mutante este x = 1/(2N) , unde 2N este numrul de locusuri.
Soarta alelei e determinat doar de ans, x crescnd i descrescnd pn cnd alela e pierdut sau fixat. Probabilitatea fixrii este de u = 1/(2N) iar probabilitatea pierderii de 1-1/(2N). Daca rata de apariie a mutaiei este de v per locus i per generaie atunci numrul de mutaii ce au loc n ntreaga populaie este de 2Nv per locus i per generaie. Cum probabilitatea fixrii mutaiei este de 1/(2N) rata de substituie per locus i per generaie (a) este de : a = Nv x1/2N=v Deci rata de substituie per locus i per generaie este egal cu rata mutaiei. Aceasta regul simpl a fost descoperit n 1968 de ctre Kimura. n cazul mutaiilor avantajoase selective situaia este alta, fitnesul indivizilor fiind de 1 n cazul genotipului AA, 1+s n cazul genotipului Aa i 1+2s n cazul genotipului aa (s este numit coeficient de selecie i este pozitiv pentru mutaiile folositoare i negativ pentru cele duntoare). Probabilitatea fixrii mutaiilor avantajoase u 2s pentru populaii mari unde Ns 1, deci rata de fixare a mutaiilor avantajoase este a = 2Nv x 2s = 4Nsv Acest exemplu este totui un pic exagerat deoarece odat ce o protein are o funcie stabilit majoritatea mutaiilor vor fi duntoare sau neutre i foarte puine vor amplifica activitatea proteinei. Mai multe studii au artat c rata mutaiilor pozitive este mult mai mic ca a celor neutre. Marea majoritate a mutaiilor sunt ns duntoare, ele fiind rapid eliminate din populaie, ele necontribuind deci la substituiile de aminoacizi i nefiind fixate. Mutaiile puin duntoare se comport altfel ns. Ele pot fi totui fixate ntr-o populaie cu o mic probabilitate n special n populaii mici. Totui o acumulare continu de mutaii puin duntoare i-ar putea deteriora structura i funcia. Deci ca mutaiile uor duntoare s fie fixate trebuie s se acumuleze i mutaii uor benefice pentru ca proteina s-i pstreze funcia. n acest caz rata medie de acumulare a mutaiilor poate deveni mai mult sau mai puin constant (pe termen lung bineneles). n unele cazuri heterozigoii (Aa) pentru o nou mutaie pot avea un fitnes crescut fa de oricare dintre homozigoi (AA sau aa) acest fenomen fiind cunoscut sub denumirea de selece supradominant. n acest caz fitnesul indivizilor AA, Aa i aa poate fi exprimat ca 1-s1, 1 i 1-s2 respectiv calculele probabilitii fixrii sunt prea complicate pentru a fi redate aici, ns cnd s1 s2 probabilitatea este de cteva ori mai mare ca cea pentru mutaiile neutre. TIPURI DE MODIFICRI LA NIVELUL MACROMOLECULEI DE ADN Exist patru tipuri principale de modificri care au loc n macromolecula de ADN: 1. nlocuirea nucleotidelor, 2. deleia, 3. adiia i 4. inversia acestora. Adiia, deleia i inversia se pot produce pentru una sau mai multe nucleotide. Adiia i deleia pot modifica secvenele cadrelor deschise de citire, poriuni numite ORF (open reading frame), caz n care, sunt numite frameshift mutations (figura).
nlocuirea nucleotidelor poate fi mprit n dou clase: tranziii i transversii. Tranziiile reprezint nlocuirea unei baze azotate purinice (adenin sau guanin) cu alt baz purinic sau a unei baze azotate pirimidinice (timin sau citozin) cu o alt pirimidin (A G; T C) Substituia nucleotidelor purinice cu nucleotide pirimidinice sau invers, poart numele de transversii (AT; GC; AC; TG). n majoritatea secvenelor de ADN, s-a constatat c substituia nucleotidelor prin tranziii, intervine mai frecvent dect substituia prin transversii (Fitch, 1967; Gojobori et al., 1982; Kocher et al., 1991). Modificrile evolutive de la nivelul macromoleculelor de ADN sunt mai complicate dect cele din secvenele proteice, deoarece exist mai multe tipuri de ADN cum ar fi regiunile codificatoare, regiuni necodificatoare, exoni, introni, regiuni terminale, regiuni repetitive i secvene de inserie. Modificrile produse de mutaii la nivelul ADN variaz n funcie de zona n care apar. Chiar dac lum n considerare o singur regiune codificatoare, modelul substituiei nucleotidelor n prima, a doua i a treia poziie a unui codon, este diferit. Mai mult, unele regiuni se afl sub aciunea seleciei naturale, mai mult dect altele, acest fapt contribuind la variaii ale modelului de evoluie ale diferitelor regiuni ale ADN.
Modul de apariie al frameshift mutaiilor
DIFERENE NUCLEOTIDICE NTRE SECVENE Cnd dou secvene de ADN provin din aceeai secven ancestral, evolueaz divergent gradual, datorit substituiei nucleotidice. Cea mai simpl msur a divergenei dintre secvene, este proporia siturilor nucleotidice care difer ntre dou nucleotide sau distana p pentru secvenele nucleotidice.
n cazul substituiei aminoacizilor, distana p ne d o estimare a numrului de nucleotide substituite per site, cnd secvenele au origine apropiat. Cnd p are valoare mare, ne d o subestimare numeric, deoarece nu ine cont de mutaiile revers i de cele paralele. Pentru a estima numrul substituiilor nucleotidice, este necesar s folosim un model matematic al substituiilor nucleotidice. Din acest motiv, a fost constituit o matri a ratei substituiilor nucleotidice (Tabelul) (Nei i Kumar, 2000). T C G A T C G Jukes-Cantor model HKY model A gT gC agG T gA agC gG C gA agT gG G agA gT gC Kimura model Tamura-Nei model A gT gC a1gG T gA a2gC gG C gA a2gT gG G a1gA gT gC Equal-input model General reversible model A agT agC agG agT bgC cgG T agA agC agG agA dgC egG C agA agT agG bgA dgT fgG G agA agT agC cgA egT fgC Tamura model Unrestricted model A 2 1 1 a12 a13 a14 T 2 1 1 a21 a23 a24 C 2 2 1 a31 a32 a34 G 2 2 1 a41 a42 a43 Not: un element (eij) care intr n una din matricile anterioare de substituie nucleotidic va nlocui nucleotida din rndul i cu nucleotida din coloana j. gA, gT, gC i gG sunt frecvenele nucleotidelor. 1 = gG + gC; 2 = gA + gT. MODELUL JUKES CANTOR Cel mai simplu model de substituie nucleotidic este cel propus de Jukes i Cantor n 1969. Acest model presupune c substituia nucleotidic intervine n orice site cu frecven egal, astfel nct, o nucleotid este nlocuit cu una din celelalte trei nucleotide rmase cu o probabilitate pe an (sau o alt unitate de timp). Mai mult, probabilitatea de nlocuire a unei nucleotide cu oricare din celelalte trei, este r=3, unde r reprezint probabilitatea de nlocuire pe site pe an. Dac lum n considerare dou secvene X i Y care au evoluat divergent dintr-o secven comun ancestral, cu t ani n urm. Dac notm cu qt nucleotidele comune dintre cele dou secvene i cu pt (pt=1 qt) proporia nucleotidelor diferite, procentul nucleotidelor comune qt+1 n timpul t+1 (msurat n ani), poate fi obinut astfel n primul rnd, trebuie s precizm c acest site care prezint nucleotide comune pentru X i Y n timpul t, va rmne neschimbat n timp, adic n t+1, cu o probabilitate (1-r)2 sau aproximativ 1- 2r, deoarece r reprezint o cantitate foarte mic astfel nct r2 poate fi neglijat. n plus, un site care prezint nucleotide diferite n timpul t, va avea aceleai nucleotide n timpul t+1 cu A
o probabilitate 2r/3. Aceast probabilitate este obinut innd cont de faptul c n momentul n care secvenele X i Z au nucleotidele i respectiv j, n timpul t, ele devin identice dac i n X se schimb cu j, dar j din Y rmne neschimbat sau dac j din Y este nlocuit cu i, iar i din X rmne neschimbat. Probabilitatea de apariie a primului eveniment este de (1-r)=r(1-r)/3, deoarece i in X trebuie s fie nlocuit cu nucleotida j (cu o probabilitate mai mare dect cazul celorlalte dou nucleotide), iar nucleotida j din Y trebuie s rmn neschimbat. Probabilitatea de apariie a celui de-al doilea eveniment este de asemenea r(1-r)/3. Mai mult, probabilitatea total este 2r(1-r)/3, care devine aproximativ 2r/3 dac nu se ine cont de valoarea lui r2. n cadrul acestui model, se presupune c rata substituiei nucleotidelor este aceeai pentru fiecare pereche de nucleotide, astfel nct frecvenele ateptate ale celor patru baze azotate A, T, C i G sunt egale cu 0,25. Oricum, nefcnd nici o precizare referitoare la frecvena iniial, valoarea lui d rmne aceeai; altfel spus, nu este necesar o constan a valorii frecvenelor nucleotidice, pentru ca ecuaia ce d valoarea diferenei (d) s fie aplicabil (Rzhetsky i Nei, 1995). MODELUL KIMURA CU 2 PARAMETRI n cazul modelului lui Kimura cu 2 parametri se ea n considerare frecvena mai mare a tranziiilor fa de cea a translaiilor, rata de substituie r fiind astfel + 2 i nu 3 . MODELUL TAMURA - NEI Unul din modelele matematice ce d concluziile de ordin filogenetic prin metoda maximum likelihood (ML) (metoda probabilitii maxime), este cel al lui Hasegawa et al. (HKY) (1985). Acest model este un hibrid ntre modelul bazat pe 2 parametri al lui Kimura i modelul inputului egal i ia n considerare att tranziiile ct i transversiile, precum i valorile coninutului n GC. Oricum, formulele pentru valoarea lui d n cadrul acestui model sunt destul de complicate (Rzhetsky i Nei, 1995), astfel nct nu ar trebui s nu-l lum n considerare. n plus, acest model este inclus n modelul propus de Tamura i Nei (1993), ca un caz special, permind soluii analitice pentru valoarea distanei (d). MODELUL TAMURA Un alt model, cel al lui Tamura din 1992 e deosebit de cel al lui Kimura prin aceea c ia n considerare faptul c frecvena celor patru nucleotide nu este egal. De exemplu la Drosophila sau la Hymenoptera ADN mitocondrial are un coninut mult mai ridicat n AT dect n GC. Deci acest model este un caz particular al celui al lui Kimura pentru cazurile n care coninutul de GC este foarte nalt sau foarte redus. d = -h ln(1- P / h -Q) - (1/ 2)(1- h) ln(1- 2Q) unde h = 2 (1- ) , iar este coninutul n GC. Dac considerm un caz n care n = i un set de date ce urmeaz modelul Tamura-Nei putem vedea cum se comport diverse metode. n Figura se observ bineneles c modelul TamuraNei se ajusteaz perfect datelor iar celelalte modele subestimeaz mai mult sau mai puin numrul real de substituii. Diferenele sunt mari pentru d 0,6 , pn la d = 0,5 modelul lui Tamura dnd rezultate practic identice cu cel Tamura Nei. Modelele Tamura, Kimura2P i Jukes Cantor dau aproape acelai rezultat cu modelul Tamura Nei pentru d = 0,25, chiar i distana p devine foarte
similar cu celelalte dac p 0,1 . Deci n cazul analizrii unor specii apropiate sau a unor gene foarte conservate cum ar fi citocromoxidaza I (COI) nu este necesar utilizarea metodelor prea complexe. n acest caz este mai indicat utilizarea unei metode simple pentru c are o varian mai mic. De asemeni ca i n cazul secvenelor de aminoacizi i n cazul celor de acizi nucleici poate fi utilizat distana gamma pentru cazurile n care rata substituiilor difer substanial n cadrul aceleiai secvene. Mai uzuale sunt distanele gamma pentru modelele Jukes-Kantor, Kimura 2P i Tamura-Nei. Distanele gamma sunt mai realiste dar au o varian mai mare, din aceast cauz rezultatele obinute nu produc n mod obligatoriu rezultate mai bune n analizele filogenetice (doar dac numrul de nucleotide folosite nu este foarte mare). Pentru estimarea lungimii ramificaiilor unui arbore filogenetic distanele gamma dau de obicei rezultate mai bune.
PARTICULARITILE PRINCIPALELOR METODE TAXONOMICE Tipologia Reflectarea Uneori reflect filogenezei n clasificare Caractere luate n Cele mai calcul importante Reflectare prezent tempoului evoluiei Reflectarea Lipsete grafic a rezultatului Tipul de date Caractere separate Nivelul ierarhic Intensitatea este determinat deosebirilor de: Fenetica Nu reflect Toate prezent Fenogram Sistematica Cladistica evoluionist Este considerat Este considerat important indispensabil Unicale (apomorfe) prezent Arbore evoluionist Unicale comune (sinapomorfe) Absent Cladogram Caractere separate Prezena caracterelor i sinapomorfe. de
Matria Caractere distanelor dintre separate taxoni Numrul de Prezena deosebiri caracterelor sinapomorfe numrul deosebiri
METODE DE RECONSTRUIRE A ARBORILOR FILOGENETICI Metodele utilizate la ora actual pentru reconstruirea arborilor filogenetici pot fi mprite n 2 mari categorii: 1. Metode bazate pe distane 2.Metodele simbolic orientate
metoda MP (Maximum Parsimony) metoda ML (Maximum Likelyhood) Reconstrucia unui arbore filogenetic este de obicei privit ca deducerea statistic a adevratului arbore filogenetic care este necunoscut. Acest proces poate fi subprit n dou etape i anume estimarea topologiei i estimarea lungimii ramificaiilor. Cnd topologia este cunoscut estimarea lungimii ramificaiilor este relatv simpl, utilizndu-se metode ca cea a ptratelor minime sau ML. Cum pentru un numr dat de secvene exist foarte multe topologii posibile au fost dezvoltate metode de optimizare a cutrii cum ar fi cea a maximei verosimilitudini (ML) sau a evoluiei minime (ME). Pentrun secvene mici i un numr de specii mare aceste metode tind ns s dea topologii greite.
METODE BAZATE PE DISTANE n cadrul metodelor bazate pe distan sau pe matrixul de distan, acestea sunt calculate pentru toate perechile de taxoni, iar arborii filogenetici sunt construii innd cont de relaiile stabilite n jurul valorilor acestor distane. n acest caz, direciile filogenetice sunt construite considernd matrici ale distanelor n care sunt calculate distanele ntre secvene luate dou cte dou. Exist un numr foarte mare de metode prin care se pot estima direciile filogenetice pornind de la distane. UPGMA (Unweighted Pair-Group Method using Arithmetic Averages) Numele metodei provine de la unweighted pair-group method using arithmetic averages" (UPGMA). Metoda este atribuit autorilor Sokal i Michener (1958), dar s-a constatat ulterior c prezenta metod este complet diferit de cea utilizat de acetia. n schimb, algoritmul apare publicat de ctre Sneath i Sokal (1973). Un arbore construit pe baza acestei metode este numit i fenogram, deoarece a fost iniial folosit la reprezentarea gradului de similaritate fenotipic pentru un grup de specii, n cadrul taxonomiei numerice. Oricum, poate fi folosit n filogenia molecular atunci cnd rata de substituie a genelor este mai mult sau mai puin constant. n special n cazul n care frecvena genelor este folosit n reconstrucia filogenetic, acest model produce arbori destul de buni, comparativ cu alte metode bazate pe distan (Nei et al., 1983; Takezaki i Nei, 1996). n acest caz, o msur a distanei care prezint un coeficient mai mic de variaie, pare s duc la construirea de arbori cu o mai mare precizie, comparativ cu alte metode de determinare a distanei, chiar dac nu respect exact numrul de substituii genice (Takezaki i Nei, 1996). Prin UPGMA se pot construi arbori pentru evoluia n cadrul speciilor, cu toate c erori de topologie apar adesea cnd rata substituiei genelor nu este constant sau cnd numrul de gene sau nucleotide este mic. n cadrul UPGMA, un mod de msurare a distanelor evolutive, este calcularea acestora pentru toate perechile de taxoni sau secvene, valorile distanelor fiind sub forma unei matrici (Tabelul 4)
Taxon 2 3 4 5
Tabel 4 Prima matrice a distanelor (Nei i Kumar, 2000) 1 2 3 4 d12 d13 d23 d14 d24 d34 d15 d25 d35 d45
Din aceast matrice, rezult c dij reprezint distana dintre taxonii i i j. Analiza grupat (de tip clustal) a taxonilor ncepe cu perechea de taxoni ntre care exist cea mai mic distan. Presupunnd c d12 este cea mai mic distan dintre toate valorile matricei rezultate, taxonii 1 i 2 vor fi grupai pornind dintr-un ram comun localizat la distana b=d12/2. Din acestea, putem presupune c lungimea ramurilor celor doi taxoni care provin dintr-un ram comun, este aceeai. n acest caz, taxonii 1 i 2 vor fi ncadrai ntr-un singur taxon compus sau grup [u=(1-2)], iar distana dintre acest grup i un alt taxon k (k1,2) este dat de: duk=(d1k+d2k)/2. n plus, obinem o nou matrice (Tabel 5) Tabel 5 A doua matrice a distanelor (Nei i Kumar, 2000) u=(1-2) 3 4 du3 du4 d34 du5 d35 d45
Taxon 3 4 5
n continuare, presupunnd c distana du3 este cea mai mic, taxonii u i 3 vor fi ncadrai ntr-un nou taxon compus sau cluster [v=(1-2-3)], cu o lungime a ramului comun de b=du3/2=(d13+d23)/(2x2). Aceast distan ntre noul cluster (grup) creat v i fiecare dintre taxonii rmai (k), este dat de: dkv=(dk1+dk2+dk3)/3. Atunci matricea se va transforma n (Tabelul 6): Tabel 6 A treia matrice a distanelor v=(1-2-3) 4 dv4 dv5 d45
Taxon 4 5
Presupunnd c dv4 este cea mai mic valoare din matricea anterioar, putem ncadra v=(1-2-3) i 4 ca avnd un ram comun dat de b=dv4/2=(d14+d24+d34)/(3x2) Este evident c ultimul taxon care se altur celorlali trei, este 5 i ramul comun este dat de: b=(d15+d25+d35+d45)/(4x2). Este de asemenea posibil, ca n cea de-a doua matrice cea mai mic distan poate fi d45 (sau oricare alta n afar de du3). n acest caz, taxonii 4 i 5, sunt unii cu ramura comun dat de b=d45/2, crendu-se astfel un nou taxon compus, v=(4-5). Distanele ntre v i alt taxon (3 i u) sunt date de d3v=(d34+d35)/2 i duv=(d14+d15+d24+d25)/4. Presupunnd c duv
este cel mai mic, taxonii u i v sunt grupai iar taxonul 3 va fi ultimul care se va altura grupului. De asemenea, dac d3v este cel mai mic, taxonii 3 i v se vor grupa primii.
METODA NEIGHBOR JOINING (NJ) Saitou i Nei (1987) au dezvoltat o metod eficient de realizare a arborilor filogenetici, care se bazeaz pe principiul evoluiei minime (ME). Aceast metod nu examineaz toate topologiile posibile, dar n fiecare stadiu de grupare a taxonilor, se folosete un principiu de evoluie minim. Aceast metod se numete Neighbour Joining (NJ) i este privit ca o versiune simplificat a metodei ME. Cnd se folosesc 4 din 5 taxoni, metodele NJ, ME dau rezultate identice (Saitou i Nei, 1987). Exist o oarecare asemnare ntre NJ i metoda adiional de realizare a arborilor, care d att topologia ct i lungimea ramurii simultan. Unul dintre conceptele importante n aceast metod, este reprezentat de neighbors, definii ca doi taxoni conectai printr-un singur nod ntr-un arbore fr punct de origine. De exemplu, taxonii 1 i 2 din arborele prezentat n figura A, sunt considerai neighbors (vecini), pentru c sunt conectai doar prin nodul A. Similar taxonii 5 i 6 sunt considerai neighbors, dar toate celelalte perechi nu. Cu toate acestea dac se combin taxonii 1 i 2 i se consider ca fiind un singur taxon, acesta, (1-2) i taxonul 3 sunt acum neighbors. Este posibil definirea topologiei unui arbore prin alturri succesive ale taxonilor vecini (nj) i producerea unor noi perechi de taxoni vecini. De exemplu, topologia arborelui din Figura A poate fi descris prin urmtoarele perechi de taxoni vecini (neighbors): (1, 2), (5, 6), (1 2, 3) i (1 2 3,4). Astfel, prin gsirea acestor perechi de taxoni vecini, se poate obine topologia arborelui.
Fig. A. Arborele filogenetic cu lungime tiut a ramurilor, pentru ase secvene Construirea unui arbore prin intermediul acestei metode, ncepe cu arborele n form de stea care este produs, pe baza presupunerii c nu exist o grupare a taxonilor (Figura B). n practic, aceast presupunere este n general incorect. Astfel, dac se estimeaz lungimea ramului unui arbore n form de stea i se calculeaz suma tuturor ramurilor (S0), aceast sum ar trebui s fie mai mare dect suma (SF) pentru arborele de tip NJ final. Dac se elimin taxonii vecini 1 i 2 din cadrul arborelui prezentat n Figura C, suma (S12) a tuturor lungimilor ramurilor, trebuie s fie mai mic dect S0, cu toate c este posibil s fie mai mare dect SF. n practic, deoarece nu se tie exact care perechi de taxoni sunt vecini, se consider toate perechile de taxoni, ca poteniale perechi de taxoni vecini i se calculeaz suma lungimilor ramurilor (Sij) pentru taxonii i i j,
utiliznd o topologie similar cu cea din Figura A, se pot alege taxonii i i j care au cea mai mic valoare pentru Sij Desigur, valorile distanelor sunt subiectul erorilor stochastice, astfel nct taxonii vecini alei n acest mod, nu sunt ntotdeauna adevraii taxoni vecini. O dat identificat o pereche de taxoni vecini, aceti sunt ncadrai ntr-un taxon compus, procedura repetndu-se pn la producerea arborelui final
Fig.B. Arbore filogenetic n form de stea
Separarea primilor taxoni vecini (1 i 2),
Aceste valori sunt determinate prin metoda ptratului minim, pentru actuala topologie a arborelui. Urmtorul pas, este calcularea distanei ntre noul nod A i taxonii rmai (k; 3km)
Separarea nodului A de restul taxonilor
Separarea taxonilor 5 i 6 prin formarea nodului B
Separarea taxonului 3 prin apariia nodului C
Separarea taxonului 4 prin apariia nodului D Dac se calculeaz toate distanele pe baza acestei valori, vom obine o nou matrice de tipul (m-1)(m-1). Pentru aceast nou matrice, putem calcula o nou sum Sij, care va fi notat cu Sij`, deoarece nu include lungimea ramurilor externe pentru prima pereche de taxoni vecini identificai, astfel nct apare ca fiind mai mic dect suma total (Sij) a lungimii braelor la acest nivel al construciei arborelui filogenetic. Pentru gsirea unei noi perechi de taxoni vecini, se ia n considerare din nou perechea cu cea mai mic valoare Sij. Un nou nod B poate fi creat pentru aceast nou pereche de taxoni i o nou valoare a matricei distanei (m-2)(m-2) este calculat. Aceast procedur se repet, pn cnd toi taxonii sunt grupai ntr-un singur arbore fr punct de origine (unrooted tree) de tip neighbour joining
Justificarea principiului evolutiv prin rezultatele altor tiine ncepnd cu primele etape de dezvoltare a concepiei filogenetice actuale, dovezile istoriconaturale luate n consideraie pentru a demonstra procesul filogenetic au servit pentru nelegerea mai profund a particularitilor procesului evolutiv. Dovezile evoluiei filogenetice sunt strns intercalate cu metodele de studiu a proceselor filogenetice. Structura tuturor organismelor vii este bazat pe structuri complexe macromoleculare de natur proteic. Moleculele de ADN care conin codul informaiei ereditare la toate formele vii (cu excepia bacteriilor) sunt strns legate structural cu moleculele proteice, formnd nucleoproteidele. Unitatea structurii chimice a plantelor i animalelor este confirmat de asemnarea principial a structurii clorofilei la plante, hemoglobinei la animalele vertebrate i a hemocianinei la animalele nevertebrate. Procesele biochimice de baz de asemenea sunt unice pentru ntreaga lume organic: oxidarea, glicoliza, descompunerea acizilor grai, transportul substanelor specifice prin membranele biologice . Pentru organismele din diferite grupe sistematice este caracteristic participarea enzimelor cu o structur foarte apropiat n reaciile biochimice de acelai tip. Studiul structurii
biochimice comune, precum i a particularitilor metabolismului la diferite grupuri de vieuitoare a dus la concluzia despre caracterul biochimic universal al vieii pe Pmnt, ceea ce este de asemenea i o dovad a unitii originii vieii. Originalitatea florei i faunei diferitor regiuni biogeografice se explic prin istoria formrii scoarei Pmntului, istoria geologic a mrilor i oceanelor. Conturul i legturile dintre continente s-au modificat de multe ori pe parcursul istoriei planetei. Izolarea profund i de lung durat a unor regiuni de uscat au dus la apariia florei i faunei unice, care se deosebete esenial de flora i fauna celorlalte regiuni ale Terrei. Apariia unor legturi ntre continente au dus la interptrunderea elementelor complexelor floristice i faunistice vecine. Astfel, marsupialele i cazuarii sunt elemente caracteristice faunei australiene; leneii,tatu, hoachinii celei sud americane, iar cactuii, bromelia i canele sunt caracteristice florei neotropicelor. Particularitile morfologice comune pot fi nelese dor cu condiia acceptrii unitii originii grupelor care posed aceste caractere. Speciile actuale prezint doar ramurile terminale ale arborelui filogenetic, trunchiul i ramurile cruia au disprut n trecut. Speciile nrudite formeaz genuri, genurile familii, familiile ordine .a.m.d. ntre aceti taxoni n natur exist decalaje din cauza lipsei tranziiilor lente. Faptul, c n baza unor complexe de date sistematicienii reuesc s stabileasc cu un grad nalt de veridicitate subordonarea ierarhic a diferitor taxoni (specia se supune genului, genul- familiei .a.m.d.) poate fi explicat doar prin existena real a legturilor evoluioniste ntre diferite grupe prin originea unora de la altele, a celor tinere de la cele btrne. Nivelul sistematic al grupei este determinat n mare msur de numrul de subdiviziuni fiice. Evidenierea genurilor, familiilor, ordinelor, claselor prezint o necesitate obiectiv ci nu se face de dragul comoditii sistematicii. Genetica i selecia prezint dovezi elocvente ale procesului evolutiv. Toate rasele de cini au aprut n rezultatul seleciei naturale din formele slbatice strmoeti. Acelai lucru poate fi spus despre rasele de vite, psri .a.. Sun cunoscute rezultatele geneticii i seleciei n crearea noilor forme de plante: n mod experimental au fost obinute nu numai soiuri i subspecii noi, dar chiar specii, i chiar genuri noi de plante. Un rol deosebit pentru teoria evoluiei filogenetice o are dezvoltarea geneticii populaionale, care permite de a elucida mecanismele, care duc la schimbarea ireversibil a frecvenei anumitor gene n populaie. Adugnd la cele spuse numrul enorm i n permanent cretere a dovezilor paleontologice ale procesului de dezvoltare istoric a vieii, putem concluziona, c de fapt avem nevoie nu att de dovezi ale evoluiei, ci de stabilirea mecanismelor concrete, care asigur procesele evolutive, precum i a particularitile acestui proces la diferite niveluri de organizare a materiei.
Principalele metode de studiu a procesului evolutiv Metodele paleontologice. n linii generale, toate metodele aplicate n paleontologie pot fi considerate ca metode, care asigur studiul procesului evolutiv. n continuare vor fi prezentate doar unele dintre acestea.
Formele fosile de tranziie forme de organisme, care combin caracterele grupelor vechi i a grupelor noi. Cutarea i descrierea unor asemenea forme asigur reconstruirea filogeniei unor grupe separate. Un reprezentant al formelor de trecere este Ichtyostega care permite de a sesiza legtura ntre peti i vertebratele terestre. Unii dintre reprezentanii strvechi ai vertebratelor terestre din grupul stegocefalilor deasemea au pstrat unele caractere ale petilor. O form de trecere de la reptile la psri sunt i Archaeopteryx . Acestea au pstrat coada lung cu vertebre neconcrescute, coaste ventrale i dini ca la reptile, dar acelai timp sunt psri adevrate cu corpul acoperit de pene, membrele anterioare transformate n aripi. Un alt exemplu din acest ir este reptila Lycaenops din grupa terapsidelor. Dezvoltarea osului mare dental, boltei osoase secundare, tipic pentru mamifere, diferenierea dinilor i alte caractere fac aceste animale asemntoare cu mamiferele rpitoare strvechi. Dar din punct de vedere al majoritii caracterelor i a modului de via acestea erau de fapt reptile adevrate. irurile paleontologice - iruri de forme fosile, legate ntre ele n procesul evolutiv, care reflect evoluia filogenetic. n dependen de numrul de forme de tranziie gsite irurile pot fi mai mult ori mai puin detaliate, dar n mod obligatoriu aceste forme trebuie s se caracterizeze prin asemnarea nu numai general, dar i detaliat dup anumite caractere, i deci, care au legturi filogenetice evidente. Asemenea iruri sunt deosebit de interesante, cnd formele lor mai noi sunt de fapt organisme, care vieuiesc n prezent pe Pmnt, fapt ce face mai accesibil nelegerii cauzele unei asemenea analize profunde a istoriei filogenetice a grupei date, veriga final a creia este convieuitorul nostru, iar formele tranziionale fosile permit restabilirea filogeniei ei. Cele mai cunoscute n prezent iruri se refer la cai, rinoceri i elefani. Analiza irului evolutiv al cailor a fost iniiat n lucrrile lui Kovalevski V.O. (1842-1883). n baza analizei irului cailor se vede succesivitatea procesului evolutiv, care confer formelor strvechi caractere tot mai apropiate i mai apropiate formelor cailor contemporani.. La examinarea comparat a eohipusului i calului contemporan este foarte greu s depistezi caracterele de asemnare ntre aceste forme nrudite , dar existena numeroaselor forme succesive de tranziie demonstreaz elocvent aceast nrudire. Succesivitatea formelor fosile. irurile paleontologice se alctuiesc n baza analizei unor constatri dispersate, care se refer la diferite regiuni geografice. E clar c unele forme lipsesc din studiu din cauza ca materialul corespunztor nu a fost depistate. Paleontologia contemporan ofer nc o dovad incontestabil a proceselor evolutive ce se nregistreaz n anumite grupuri. n anumite condiii favorabile n acelai loc se pstreaz n form de fosile toate formele disprute. Analiza straturilor acestor depuneri permite reconstruirea succesiunii reale a apariiei acestor forme. Prioritatea acestei metode const n aceea, c ea ofer posibilitatea aprecierii reale a vitezei procesului evolutiv.
Studiul evoluiei ecosistemelor. Aplicarea metodelor moderne de cercetare n paleontologie permite de a obine date noi, tot mai complete despre istoria vieii pe Terra. Astfel, prin aplicarea metodei de dizolvare a rocilor nconjurtoare, este posibil de a restabili cele mai fine structuri fosile, inclusiv a microorganismelor. Aplicarea metodelor paleomagnetice, paleochimice, de radioautografie .a. permit de a obine date veridice despre clim i condiiile de existen din diferite perioade: despre salinitatea oceanului, temperatura apei i a aerului, componena atmosferei .a. Sunt depistate tot mai multe forme fosile. Toate aceste date completeaz golurile n cunotinele actuale, dar cu parere de ru nc nu le completeaz pe toate. n baza noilor realizri ale tiinei devine posibil reconstrucia succesiunii ecosistemelor. Asemenea reconstruiri succesive a tabloului anumitor regiuni ale planetei sunt nite metode integrate foarte puternice n stabilirea direciei procesului evolutiv pe Pmnt. Fiind metodele de baz pentru studiul macroevoluiei (care decurge n scara de sute de mii milioane de ani), metodele paleontologice sunt puin informative pentru studiul microevoluiei (care decurge n scara de sute mii de ani).
Metodele biogeografice. Metodele biogeografice permit de a analiza mersul evoluiei la diferite nivele. Compararea florei i faunei. Despre procesul evolutiv la nivelul apariiei noilor flore i faune se poate de vorbit n contextul evoluiei continentelor pe planet. La sfritul perioadei triasice Pangea s-a desfcut n dou supercontinente: de nord Lavrasia i de sud Gondvana. Gondvana la rndul su s-a desfcut i a format continentul African-Sudamerican; Australia cu Antarctida; i o insul enorm, care mai trziu a aderat la Asia formnd Indostanul. a sfritul perioadei Jurasice Lavrasia se dezmembreaz i formeaz America de Nord i Euroasia, iar America de Sur cu Australia i Madagascarul mai formeaz nc un monolit . Urmele acestei uniti se pstreaz i pn n zilele noastre n componena faunei. La acestea se refer iguanele Madagascarului i Americii de Sud, somnii i petii haracini din America de sud i Africa. Cea mai apropiat rudenie a Broatei africane cu gheare este broasca fagure, care poart pe spate larvele n dezvoltare. n acelai timp broasca paradoxal sud american, mormolocul creia ca i mormolocul broatei de pmnt este mai mare ca forma matur, este legat prin legturi mult mai slab pronunate cu broasca rioas australian de pustiu. Acest lucru este clar reieind din faptul, c Antarctida i Australia au reuit deja s se separeu (iar n Australia au reuit s ptrund strmoii marsupialelor, care semnau cu oposumul american o specie actual de animale marsupiale. La sfritul perioadei cretacice - nceputul neogenului conturul continentelor se apropie de cel actual, apare Oceanul Atlantic, care iniial era nu prea lat, astfel c pe trunchiuri plutitoare din Africa n America de Sud se deplasau maimuele primitive i porcii cu epi. ncepe s apar fisura ntre Australia i Antarctica. Madagascarul se rupe de la Africa i devine o rezervaie a formelor strvechi. Indostanul se mica spre nord ca n sfrit s se ciocneasc cu Asia, s acopere marea Tetis i s formeze munii Himalaia. Laurasia rmne nc incomplet separat, iar schimbul de forme ntre Lumea Veche i cea Nou continu.
Vom analiza cteva exemple de asemnare i deosebire a populaiilor diferitor teritorii din punct de vedere evoluionist. Fauna mamiferelor Europei de Vest se pstreaz foarte asemntoare cu cea de peste Ural a Asiei de Nord. Acest lucru se explic prin faptul, c pe teritoriul enorm al Euroasiei nu exist bariere de netrecut pentru formele mari i medii de mamifere. Fauna mamiferelor Africii de nord este foarte asemntoare cu cea a Asiei de Nord, iar deosebirile nu depesc de regul nivelul de gen. Pentru fauna Americii de nord ca i pentru cea a Euroasiei de Nord sunt caracteristice asemenea forme ca elanii, jderii, nurca, volverina, urii polari, pika, veveria zburtoare, marmota, popndul .a. Aceast asemnare se explic prin faptul, c pn nu demult (cel mult un milion de ani) ntre Euroasia i America de Nord a existat un pod Beringhia. La compararea faunei mamiferelor Americii de Nord i celei de Sud, nectnd la apropirea lor teritorial, deosebirile sunt enorme, la nivel de genuri, n calitate de endemice pentru America de sud fiind mai mult de 80%. Numai aici locuiesc leneii (Bradypodidae), furnicarii (Myrmecophagidae), tatu (dasypodidae). Nu mai puin se deosebete i ornitofauna Americii de Sud. O asemenea originalitate a faunei acestui continent se explic prin faptul, c mai multe zeci de milioane de ani America de Sud a fost complet izolat de alte pri ale uscatului. Istmul Panama, care a aprut de cteva ori pe parcursul istoriei era prea ngust pentru a asigura un schimb intens de forme ntre continente. Aceeai situaie s-a creat i n Australia, care timp de 120 milioane de ani nu a avut ci de comunicare cu alte continente. n acest teritoriu s-a dezvoltat independent, fr implicarea faunei din alte regiuni a mamiferelor cu cloac i a marsupialelor. Istoria dezvoltrii faunei australiene numr n general 4 etape. Prima etap este ptrunderea (sau dezvoltarea pe loc a Marsupialelor i Monotrematelor, care au dat natere varietii actuale de specii. A doua etap s-a nregistrat n timpul scderii eseniale a nivelului Oceanului planetar, cnd de pe iruri de insule n Australia au ptruns multe specii de roztoare. A treia etap const n introducerea cinelui slbatic Dingo, care mai degrab a nimerit aici cu omul cu cteva zeci de mii de ani n urm i n sfrit, n epoca contemporan pe continent au fost introduse cu succes specii de iepuri, cerbi i alte mamifere. O analiz de tipul celei efectuate mai sus pentru unele specii de mamifere poate fi realizat pentru specii de plante ori alte animale. Metoda dat ofer posibilitatea de a estima, ct de strnse sunt legturile ntre biodiversitatea unor anumite regiuni ale planetei i istoria dezvoltrii acestor regiuni. Particularitile de rspndire a formelor apropiate . Pn n prezent n unele ruri ale Angliei i Irlandei se ntlnesc specii de pete alb de ap dulce din genul Coregonus, care n trecut era rspndit pe ntregul teritoriu al Europei de Nord-Vest. Pe locul Mri Irlandei era un es larg cu rezervoare enorme cu ap dulce. Coborrea nivelului Europei a dus la ptrunderea apei marinei n bazinele esului. Petele alb de ap dulce s-a retras n rurile, care au fost izolate prin masivele de ap de mare i au evoluat acolo n specii originale. Acest exemplu demonstreaz, cum n baza datelor zoogeografice poate fi reconstruit procesul evolutiv i pot fi obinute date despre viteza real a acestui proces. O asemenea analiz poate de asemenea s ne ajute n stabilirea centrelor de origine a anumitor grupe, prin evidenierea locurilor concentrrilor numerice i de biodiversitate.
Formele insulare. Individualitatea formelor insulare este cu att mai mare cu ct mai veche este sciziunea dintre insul i restul uscatului. Astfel insulele Britanice care s-au separat recent, se caracterizeaz printr-un numr mic de specii endemice. n acelai timp Fauna Madagascarului care s-a separat de foarte mult vreme de restul uscatului numr 32 genuri endemice de mamifere din cele 36 prezente pe insul, iar din ornitofaun endemice sunt jumtate din genurile prezente pe insul. Analiza detaliat a faunei insulare permite de a stabili cile evoluiei grupelor de specii nrudite. Un exemplu clasic n acest sens este cel prezentat de Darvin cintezele insulelor Galapagos. Vrsta insulelor este de cel mult cteva milioane de ani. Careva strmo al cintezelor actuale a nimerit aici cu mult naintea altor paseriforme. Acest strmo a evoluat dnd natere variatelor forme actuale (13 specii). La categoria formelor insulare se refer i speciile de peter. n petere se ntlnesc forme, care explic cile evolutive recente. Astfel n Petera Mamuilor din America de Nord se ntlnete petele orb de peter (Amblyopsis spelaea) care se ntlnete foarte larg n bazinele acvatice ale acestei regiuni. Spre deosebire de acest exemplu formele nrudite ale proteului ( Proteus anguinus) din apele subterane din Slovenia sunt amfibienii din genul Necturus din America de Nord. Rspndirea discontinu. n istoria dezvoltrii planetei sunt dese cazurile, cnd o specie rspndit pretutindeni dispare din marea parte a arealului n rezultatul modificrii condiiilor i rmne ntr-un numr mic de regiuni mici, unde s-au pstrat condiii similare celor iniiale. Un asemenea exemplu este fauna Alpilor un complex de specii rspndite n era glaciar. n prezent aceste specii populeaz doar mici regiuni alpine de altitudine mare. Un alt exemplu este lcrimioara (Convallaria majalis) care populeaz yona pdurilor europene. n Caucaz se ntlnete o specie foarte apropiat Convallaria transcaucasica, precum i n Extremul orient - Convallaria keiskei. n trecu ntreaga euroasie era populat de o form de lcrimioar, care n rezultatul naintrii ghearilor a fost localizat n anumite regiuni i s-a dezvoltat formnd specii diferite. Examinarea a unor asemenea situaii permit de a cunoate particularitile de dezvoltare a vieii n diferite regiuni ale planetei. Formele relicte. Despre dezvoltarea filogenetic a vieii pe Pmnt vorbesc foarte elocvent formele relicte care au ajuns pn n zilele noastre din trecutul ndeprtat al planetei. Relictele sunt specii ori grupuri mici de specii care posed un ir de caractere tipice pentru formele demult disprute. Una din cele mai cunoscute forme relicte este gateria (Sphenodon punctatus), care posed caracterele reptilelor disprute cu zeci de milioane de ani n urm i n prezent poate fi ntlnit doar n unele regiuni ale Noii Zelande. Un alt relict este petele crosopterigian Latimeria caracteristic devonului i care a ajuns pn n zilele noastre n unele regiuni ale apelor teritoriale ale Africii de est. ntre palte relict este specia de Ginkgo biloga, care n prezent se ntlnete doar n China i Japonia.
Astfel, metodele biogeografice permit de a studia att procesele macroevoluiei, ct i pe cele ale microevoluiei. n legtur cu aplicarea lor pentru studiul procesului microevolutiv au fost introduse chiar discipline noi, aa ca biogeografia dinamic i geno(feno)geografia.
Metodele morfologice. Aceste metode se bazeaz pe afirmaia, c o asemnare morfologic profund poate indica asupra unui grad nalt de rudenie ntre diferite forme. Omologia organelor . Organele cu un plan comun de structur, care provin din acelei germen, care sunt n relaie asemntoare cu alte organe i care ndeplinesc att funciile iniiale, ct i cele secundare sunt denumite omoloage. Stabilirea gradului de omologie ntre diferite organe ale unor forme permit de a judeca despre gradul de nrudire filogenetic a acestor forme. Este foarte important de a delimita noiunea de organe analoage, care se caracterizeaz doar prin asemnarea exterioar a anumitor organe, ceea ce este o dovad a realizrii aceleiai funcii i nu poate fi o dovad de rudenie ntre formele studiate. Organele rudimentare i atavismele. Organe rudimentare sunt acelea care apar n faza embrionar sau n faza de cretere a organismului, iar la maturitate dispar sau degenereaz (cum ar fi timusul). De aici evoluionitii au tras concluzia c aceste organe ar fi nefolositoare i existena lor nu ar putea fi explicat dect dac presupunem c acestea ar fi nite rmie din stadiile anterioare ale evoluiei speciei respective. La o analiz mai atent se constat totui c aceste organe sunt nefolositoare numai n faza adult, pe cnd n timpul creterii au un rol important n organism, iar faptul c apar numai atunci cnd este nevoie de ele iar apoi dispar nu face dect s confirme existena finalitii i s contrazic teoria evoluionist, deoarece dac aceste organe ar fi rmase de la specii anterioare atunci nu ar disprea la maturitate. Spre deosebire de organele rudimentare care exist la toi indivizii, organele atavice apar numai uneori, constituind nite anomalii sau monstruoziti. Din faptul c unele din aceste malformaii existente la om, de exemplu, prezint asemnri cu structuri sau organe existente la unele animale, evoluionitii trag concluzia c omul descinde din animalele respective i c, dat fiind inutilitatea acestor organe la om, ele ar fi i o prob mpotriva finalitii. Ei uit ns s precizeze c ntre malformaii exist i foarte multe care nu prezint nici o asemnare cu vre-o structur existent la animale. n plus, dintr-o asemnare exterioar a unor organe sau esuturi, nu rezult n mod logic faptul c omul ar descinde din animalul respectiv (cu att mai mult cu ct unele malformaii se aseamn cu animale pe care nici evoluionitii nu le consider strmoi ai omului). n ceea ce privete finalitatea, organele atavice nu dovedesc nimic, deoarece reprezint stri patologice. Unii evoluioniti, dat fiind evidena faptului c toate vieuitoarele posed exact acele organe care le sunt necesare n mediul n care triesc, accept ntr-o oarecare msur ideea de finalitate, numai c pun carul naintea boilor afirmnd c funcia creeaz organul. Dar aceast afirmaie nu este nici logic (cum ar putea exista, de exemplu, funcia vederii nainte de apariia
ochilor?) i vine n contradicie cu observaiile experimentale care arat c n timpul dezvoltrii unui organism, organele ncep s se formeze cu mult nainte de a ndeplini vreo funcie (de exemplu, copilul nou nscut are deja picioare, dar abia mai trziu nva s mearg). Aadar nu funcia creeaz organul, ci organul a fost proiectat pentru a ndeplini o anumit funcie, proiectul anticipnd funcia pe care urmeaz s fie ndeplinit. Morfologia populaional. Metodele morfologo-populaionale permit de a depista direcia seleciei naturale dup modificarea rspndirea valorilor caracterului ntr-o populaii la diferite etape de dezvoltare a ei, sau comparativ cu alte populaii. METODE UTILIZATE N STUDIUL GENETIC AL POPULAIILOR Populaiile pot fi studiate din punct de vedere genetic cu ajutorul a dou metode: metoda descriptiv i metoda genetica. Metoda descriptiv se bazeaz pe caracterizarea fenotipic a indivizilor unei populaii, considerndu-se c fenotipul biotipurilor din populaie reprezint expresia genotipului indivizilor n strns relaie cu mediul nconjurtor. Studiind fenotipul organismelor se pot stabili unele raporturi de dependen existente ntre indivizii aceleiai colectiviti. Deoarece ntr-o populaie exist indivizi asemntori fenotipic, dar desoebii din punct de vedere genotipic, nseamn c analiza populaiei prin metoda descriptiv nu ofer o imagine real n ceea ca privete structura genetic a acesteia. Metoda genetic de studiere a populaiilor a fost posibil numai dup descoperirea legilor lui G. Mendel de segregare a caracterelor in descendenele hibride. Analiza hibridologic permite studierea formelor rezultate n urma segregrii organismelor heterozigote. Cu ajutorul metodei genetice este posibil evidenierea, din diversitatea genotipurilor unei populaii, a acelor genotipuri care corespund mai bine obiectivelor propuse de realizat n procesul de ameliorare. n analiza genetic a populaiilor hibride se folosete metoda biometric, completat cu calculul statistic al datelor, obinnd, astfel, modelul teoretic al populaiei, pornind de la o anumit situaie dat i urmrind influena exercitat de diferii factori care acioneaza asupra acesteia. Astfel de studii genetice asupra populaiilor, bazate pe mbinarea metodei biometrice cu calculul statistic, a efectuat W. Johansen (1903), care a definit att populaia, ct i gruprile componente de indivizi din cadrul populaiei, adic liniile pure. Populaiile autogame, precum i soiurile autogame, pot fi descompuse n linii pure prin alegerea descendenilor pe parcursul mai multor generaii i prin consangvinizare. De asemenea, n analiza genetic a populaiilor se studiaz frecvena genelor i a genotipurilor din populaiile naturale, dup metoda elaborat de G. Hardz i W. Weinberg (1908), care au demonstrat caracterul legic al echilibrul genetic al populaiilor.
S-ar putea să vă placă și
- AMINOACIZIDocument34 paginiAMINOACIZIlavinia crista100% (2)
- BiochimieDocument142 paginiBiochimieStefan Alexandru100% (4)
- REPRODUCEREA La PlanteDocument3 paginiREPRODUCEREA La PlanteLuminita ComanÎncă nu există evaluări
- BotanicaDocument178 paginiBotanicaMadalin Cernica100% (1)
- Amintiri Din CopilarieDocument9 paginiAmintiri Din CopilarieCerber LaurÎncă nu există evaluări
- Evolution IsmDocument78 paginiEvolution Ismfasolăi100% (2)
- FITOPATOLOGIEDocument217 paginiFITOPATOLOGIERistin Dan Marius100% (2)
- FITOPATOLOGIEDocument217 paginiFITOPATOLOGIERistin Dan Marius100% (2)
- Sistematica FanerogamelorDocument62 paginiSistematica Fanerogamelorzahariagheorghe100% (1)
- CURS 1 Microbiologia Este Stiinta Care Se Ocupa Cu Studiul MicroorganismelorDocument13 paginiCURS 1 Microbiologia Este Stiinta Care Se Ocupa Cu Studiul MicroorganismelorDoroftei AlexÎncă nu există evaluări
- Cod GeneticDocument13 paginiCod GeneticVladlena Burțev100% (1)
- Structura Celulei BacterieneDocument17 paginiStructura Celulei BacterieneAlexandra LeahuÎncă nu există evaluări
- Studiul Diviziunii MitoticeDocument7 paginiStudiul Diviziunii MitoticeEstephy DeeÎncă nu există evaluări
- LISTALUCRARIANCANEGURADocument10 paginiLISTALUCRARIANCANEGURACornel MihaiÎncă nu există evaluări
- Dreptul Copiilor La SanatateDocument5 paginiDreptul Copiilor La SanatateAna MariaÎncă nu există evaluări
- Prezentare AnticorpiDocument15 paginiPrezentare AnticorpiOanaDorofteÎncă nu există evaluări
- Legea Hardy WeinbergDocument24 paginiLegea Hardy WeinbergmamaiesitataieÎncă nu există evaluări
- v1 Bac Bio Xi Xii e 2005Document2 paginiv1 Bac Bio Xi Xii e 2005ursuceÎncă nu există evaluări
- Domenii de Aplicabilitate Si Consideratii Bioetice in Genetica Umana - Bac AnatomieDocument1 paginăDomenii de Aplicabilitate Si Consideratii Bioetice in Genetica Umana - Bac AnatomieMadalina UngureanuÎncă nu există evaluări
- Eseu ReguliDocument17 paginiEseu RegulimacoveidumitruÎncă nu există evaluări
- Tesuturi FundamentaleDocument1 paginăTesuturi FundamentalecristinachiuaruÎncă nu există evaluări
- Studiul Microbiologic Al MucegaiurilorDocument12 paginiStudiul Microbiologic Al Mucegaiurilorpopa.janosÎncă nu există evaluări
- Genetica PopulatiilorDocument15 paginiGenetica PopulatiilorMaria Hutu0% (1)
- Fizio VegetalaDocument70 paginiFizio VegetalaAlexandra CabatÎncă nu există evaluări
- Mostenirea Independenta A CaracterelorDocument7 paginiMostenirea Independenta A Caracterelorludvig_cazacÎncă nu există evaluări
- Chestionar InterdisciplinaritateaDocument1 paginăChestionar InterdisciplinaritateaTaniaPopescuÎncă nu există evaluări
- Drepturi de Autor Si Drepturi ConexeDocument73 paginiDrepturi de Autor Si Drepturi ConexemacoveidumitruÎncă nu există evaluări
- Ovazul in Republica MoldovaDocument8 paginiOvazul in Republica MoldovaLaurentiu MunteanuÎncă nu există evaluări
- Structura Celulei ProcarioteDocument4 paginiStructura Celulei ProcarioteElenaCÎncă nu există evaluări
- ImunogeneticaDocument20 paginiImunogeneticaIleana IoanaÎncă nu există evaluări
- Alte Tipuri de Reproducere În Lumea Vie.Document2 paginiAlte Tipuri de Reproducere În Lumea Vie.Geta Gug100% (2)
- Circuitul Biotic Al Materiei În Biosferă.Document4 paginiCircuitul Biotic Al Materiei În Biosferă.Christina FergusonÎncă nu există evaluări
- Biologie Vegetala Si AnimalaDocument9 paginiBiologie Vegetala Si AnimalaEmi LpÎncă nu există evaluări
- Microbiologie Regn ProtistaDocument8 paginiMicrobiologie Regn ProtistaAndreea Paun100% (1)
- MUTAȚIIDocument12 paginiMUTAȚIITalos MiriamÎncă nu există evaluări
- Aparatul Golgi GalinaDocument14 paginiAparatul Golgi GalinaIon IsacÎncă nu există evaluări
- Cariotipul Uman Normal Si Aberatiile CromozomialeDocument6 paginiCariotipul Uman Normal Si Aberatiile Cromozomialeiuzarneim100% (1)
- Organismul Plantelor SuperioareDocument1 paginăOrganismul Plantelor SuperioareDaniela SanduÎncă nu există evaluări
- Reproducerea La Gimnosperme Si AngiospermeDocument13 paginiReproducerea La Gimnosperme Si AngiospermeloreÎncă nu există evaluări
- Biologie AnimalaDocument89 paginiBiologie AnimalaOana OnciucÎncă nu există evaluări
- Procesul de Transpiratie La PlanteDocument6 paginiProcesul de Transpiratie La PlanteMARYÎncă nu există evaluări
- BacteriiDocument4 paginiBacteriiivanalexandraÎncă nu există evaluări
- CURS 1 Sistematica Taxonomia Filogenia PlantelorDocument8 paginiCURS 1 Sistematica Taxonomia Filogenia PlantelorAnnya StephhÎncă nu există evaluări
- Plan Orientativ Pentru Lucrul Individual.Document3 paginiPlan Orientativ Pentru Lucrul Individual.mariana gmgÎncă nu există evaluări
- T1 Segregarea Perechi CaractereDocument21 paginiT1 Segregarea Perechi CaractereNicoleta CirstianÎncă nu există evaluări
- Factorii Care Influenteaza Transpiratia La PlanteDocument3 paginiFactorii Care Influenteaza Transpiratia La PlanteMARYÎncă nu există evaluări
- Basidiomycota Basidiomycotina Basidiomicete BazidiomiceteDocument4 paginiBasidiomycota Basidiomycotina Basidiomicete BazidiomiceteAgafitei MarcelaÎncă nu există evaluări
- Biologie Clasa-Ix Fisa Diviziunea IndirectaDocument2 paginiBiologie Clasa-Ix Fisa Diviziunea IndirectaIoana RusÎncă nu există evaluări
- Recapitulare FotosintezaDocument1 paginăRecapitulare FotosintezaRazvan BaniaiÎncă nu există evaluări
- Alge VerziDocument1 paginăAlge VerziNicoleta DarieÎncă nu există evaluări
- Hemoglobina Si Clorofila - Doc Cami - Doc Cu PozeDocument5 paginiHemoglobina Si Clorofila - Doc Cami - Doc Cu PozeCamelia Manea50% (2)
- Adolescentul Din Ziua de AziDocument10 paginiAdolescentul Din Ziua de AziAnaigroeÎncă nu există evaluări
- Lucrul Individual 2Document3 paginiLucrul Individual 2MihaiÎncă nu există evaluări
- Disciplinele Optionale in RomaniaDocument3 paginiDisciplinele Optionale in RomaniaBianka NicoletaÎncă nu există evaluări
- Anexa C1. Ereditate - VariabilitateDocument7 paginiAnexa C1. Ereditate - VariabilitateangelicaÎncă nu există evaluări
- GraulDocument1 paginăGraulAna-Maria AlexandruÎncă nu există evaluări
- 5 Obiective OperationaleDocument2 pagini5 Obiective OperationaleAndreea PetreaÎncă nu există evaluări
- Planificare Materie Bacalaureat Biologie Vegetala Și AnimalaDocument1 paginăPlanificare Materie Bacalaureat Biologie Vegetala Și AnimalaSorin CimpeanuÎncă nu există evaluări
- Lectie Muschi Ferigi EvalDocument11 paginiLectie Muschi Ferigi EvalVinatoru Irina100% (1)
- Chimia Sistemelor Deschise BiologiceDocument12 paginiChimia Sistemelor Deschise BiologiceAnca Alexandra VighÎncă nu există evaluări
- Integrarea Numerica Metoda Dreptunghiurilor Pentru Calculul Aproximativ Al Integralei MDDocument23 paginiIntegrarea Numerica Metoda Dreptunghiurilor Pentru Calculul Aproximativ Al Integralei MDValeria Plîngău50% (2)
- Fisa de Evaluare Ereditatea Si VariabilitateaDocument2 paginiFisa de Evaluare Ereditatea Si VariabilitateaRodica Balan0% (2)
- Lanțuri Trofice Simple-Schema LecțieiDocument1 paginăLanțuri Trofice Simple-Schema LecțieicarmenÎncă nu există evaluări
- Celula EucariotaDocument16 paginiCelula EucariotaMirela ArdeleanÎncă nu există evaluări
- Micro General ADocument520 paginiMicro General Anykol1100% (1)
- AmidonulDocument8 paginiAmidonuladyilieb100% (1)
- Relatii InterspecificeDocument3 paginiRelatii InterspecificeAdrian CojocaruÎncă nu există evaluări
- Curs NevertebrateDocument123 paginiCurs NevertebrateSima CezarÎncă nu există evaluări
- Curs 4Document12 paginiCurs 4Stefi MaryÎncă nu există evaluări
- Analiza Educaţiei Narly C.Document4 paginiAnaliza Educaţiei Narly C.macoveidumitruÎncă nu există evaluări
- Protec - Ia Soiurilor de PlanteDocument24 paginiProtec - Ia Soiurilor de PlanteIon BusuiocÎncă nu există evaluări
- Toma LiteraturaDocument80 paginiToma LiteraturamacoveidumitruÎncă nu există evaluări
- Plantele Modificate Genetic Si Impactul Lor Asupra MediuluiDocument4 paginiPlantele Modificate Genetic Si Impactul Lor Asupra MediuluimacoveidumitruÎncă nu există evaluări
- 7 GeneticaDocument9 pagini7 GeneticamacoveidumitruÎncă nu există evaluări
- Identificaţi Resursele Ironiei Şi Ale Umorului În Povestirile Lui Ion Creangă Din Ciclul Moş Ion Roată Şi Faceţi Analogii Cu Trăsăturile PersonajelorDocument2 paginiIdentificaţi Resursele Ironiei Şi Ale Umorului În Povestirile Lui Ion Creangă Din Ciclul Moş Ion Roată Şi Faceţi Analogii Cu Trăsăturile PersonajelorDaria Maxim100% (1)
- Metabolismul Ac - NucleiciDocument153 paginiMetabolismul Ac - NucleicimacoveidumitruÎncă nu există evaluări
- Dragostea Este Sacrã Şi Veşnicã Şicreşte În Tãrie Şi FrumuseţeDocument3 paginiDragostea Este Sacrã Şi Veşnicã Şicreşte În Tãrie Şi FrumuseţemacoveidumitruÎncă nu există evaluări
- 3 Genetic MolecularDocument12 pagini3 Genetic MolecularMerishor MeryÎncă nu există evaluări
- Cap1-2-2 Plasmide Replicare ReglajDocument6 paginiCap1-2-2 Plasmide Replicare ReglajLarisa-DÎncă nu există evaluări
- Cap1-2-2 Plasmide Replicare ReglajDocument6 paginiCap1-2-2 Plasmide Replicare ReglajLarisa-DÎncă nu există evaluări
- 09 GeneticaDocument4 pagini09 GeneticamacoveidumitruÎncă nu există evaluări
- Genetica Microorganismelor Şi Inginerie Genetică Microbiană NoteDocument19 paginiGenetica Microorganismelor Şi Inginerie Genetică Microbiană Notecreatza20100% (4)
- Tema 1 ATZ. Originea VietiiDocument4 paginiTema 1 ATZ. Originea VietiiDonald CunninghamÎncă nu există evaluări
- Universitatea Pedagogică de StatDocument15 paginiUniversitatea Pedagogică de StatmacoveidumitruÎncă nu există evaluări
- CelulaDocument2 paginiCelulamacoveidumitruÎncă nu există evaluări
- Legea RCA Nr. 414Document20 paginiLegea RCA Nr. 414Adelina Mihaela BichirÎncă nu există evaluări