S-ar putea să vă placă și
- ARC 8 1 (Special Section) 25-32 E79 Low ResDocument8 paginiARC 8 1 (Special Section) 25-32 E79 Low ResAmelia She M UÎncă nu există evaluări
- Artigo PatiDocument11 paginiArtigo PatiProfessor Gabriel Ribeiro - UFRBÎncă nu există evaluări
- Morato - 2011 - Bothropoides Jararaca Prey - Herp - ReviewDocument2 paginiMorato - 2011 - Bothropoides Jararaca Prey - Herp - ReviewSérgio Augusto Abrahão MoratoÎncă nu există evaluări
- Motta Tavaresetal.2016 EcologyofPhyllodytesluteolusDocument12 paginiMotta Tavaresetal.2016 EcologyofPhyllodytesluteolusvanessa e bruno becaciciÎncă nu există evaluări
- Martins Etal Final1Document3 paginiMartins Etal Final1Ana Karolina MorenoÎncă nu există evaluări
- Sales Et Al 2011 - Feeding Ecology of Ameiva AmeivaDocument10 paginiSales Et Al 2011 - Feeding Ecology of Ameiva AmeivaRaul SalesÎncă nu există evaluări
- 24324-Texto Do Artigo-95554-1-10-20091119Document10 pagini24324-Texto Do Artigo-95554-1-10-20091119Thiago de AlcântaraÎncă nu există evaluări
- Trophic Niche Pseudopaludicola BolivianaDocument7 paginiTrophic Niche Pseudopaludicola BolivianaPatrick SanchesÎncă nu există evaluări
- 13 Esponjas Porifera Parque Nacional San Esteban 2010Document14 pagini13 Esponjas Porifera Parque Nacional San Esteban 2010carmenÎncă nu există evaluări
- Lizards, Snakes, and Amphisbaenians From The Quaternary Sand Dunes of The Middle Rio Sao Francisco, Bahia, BrazilDocument12 paginiLizards, Snakes, and Amphisbaenians From The Quaternary Sand Dunes of The Middle Rio Sao Francisco, Bahia, Braziluseparadown downÎncă nu există evaluări
- Influence of Sponge Morphology On The Composition of The PolychaeteDocument7 paginiInfluence of Sponge Morphology On The Composition of The PolychaetePedro Henrique ClerierÎncă nu există evaluări
- Notes On The Courtship Behavior of Aplastodiscus ADocument9 paginiNotes On The Courtship Behavior of Aplastodiscus ABeatriz BritoÎncă nu există evaluări
- HB11 - 2022 - 2 - Carvalho - Rhinella PredationDocument7 paginiHB11 - 2022 - 2 - Carvalho - Rhinella PredationWagner M. S. SampaioÎncă nu există evaluări
- 1808-9798 (2007) 2 (31:araoga) 2 0 Co 2Document9 pagini1808-9798 (2007) 2 (31:araoga) 2 0 Co 2Isabella ValenciaÎncă nu există evaluări
- Anuran Fauna, Environments and Calling Period in Espigão do Oeste, RondôniaDocument6 paginiAnuran Fauna, Environments and Calling Period in Espigão do Oeste, RondôniaDouglas LymaÎncă nu există evaluări
- Oliveira Et Al. 2019 - Lizard Assemblages On Sandy Coastal PlainsDocument12 paginiOliveira Et Al. 2019 - Lizard Assemblages On Sandy Coastal PlainsJuliane RibeiroÎncă nu există evaluări
- Maia Et Al. 2011 Diet of The Lizard Ecpleopus GaudichaudiiDocument6 paginiMaia Et Al. 2011 Diet of The Lizard Ecpleopus GaudichaudiiThiago Maia CarneiroÎncă nu există evaluări
- Ambientes e Temporada de Vocalização Da Anurofauna No Município de Espigão Do Oeste, Rondônia, Sudoeste Da Amazônia - Brasil (Amphibia: Anura)Document6 paginiAmbientes e Temporada de Vocalização Da Anurofauna No Município de Espigão Do Oeste, Rondônia, Sudoeste Da Amazônia - Brasil (Amphibia: Anura)Paulo Sérgio BernardeÎncă nu există evaluări
- Breeding Activity of Scinax Centralis (Anura, Hylidae) in Central BrazilDocument5 paginiBreeding Activity of Scinax Centralis (Anura, Hylidae) in Central BrazilSheila AndradeÎncă nu există evaluări
- Weldele Et Al Gambusia Vittata Final Versionpdf 1Document23 paginiWeldele Et Al Gambusia Vittata Final Versionpdf 1api-326851466Încă nu există evaluări
- Diversidad Alfa y Beta de Los Artrópodos en Diferentes Ambientes Del Parque Nacional Los Cardones, Salta (Argentina)Document14 paginiDiversidad Alfa y Beta de Los Artrópodos en Diferentes Ambientes Del Parque Nacional Los Cardones, Salta (Argentina)Pedro Jorge Jiménez PradoÎncă nu există evaluări
- Epibiont Occurrence On Gastropod Shells Used by The Hermit Crab Loxopagurus Loxochelis (Anomura: Diogenidae) On The Northern Coast of São Paulo, BrazilDocument6 paginiEpibiont Occurrence On Gastropod Shells Used by The Hermit Crab Loxopagurus Loxochelis (Anomura: Diogenidae) On The Northern Coast of São Paulo, Brazilonurb90Încă nu există evaluări
- Vansluys 2007Document7 paginiVansluys 2007samanthavieira.s39Încă nu există evaluări
- 1 PBDocument11 pagini1 PBjose.1711717678Încă nu există evaluări
- Avifauna Asociada A Un Tramo Urbano Del RíoDocument10 paginiAvifauna Asociada A Un Tramo Urbano Del RíoMartinOcañaÎncă nu există evaluări
- Marangoni Et Al. 2011 - NWJZDocument9 paginiMarangoni Et Al. 2011 - NWJZRodrigo CajadeÎncă nu există evaluări
- Jurnal CephalochordataDocument3 paginiJurnal CephalochordataAnna KareninaÎncă nu există evaluări
- Leaf Morphological Strategies of Seedlings and Saplings of Rhizophora Mangle (Rhizophoraceae), Laguncularia Racemosa (Combretaceae) andDocument14 paginiLeaf Morphological Strategies of Seedlings and Saplings of Rhizophora Mangle (Rhizophoraceae), Laguncularia Racemosa (Combretaceae) andAndressa PelozoÎncă nu există evaluări
- 1676 0603 BN 13 03 93Document9 pagini1676 0603 BN 13 03 93alexandra gonzalez castilloÎncă nu există evaluări
- Article: Biota Neotropica 21 (1) : E20200985, 2021Document6 paginiArticle: Biota Neotropica 21 (1) : E20200985, 2021Camila MilÎncă nu există evaluări
- TMP 52 B6Document8 paginiTMP 52 B6FrontiersÎncă nu există evaluări
- Population dynamics, reproductive biology and ichthyoplankton of Cetengraulis edentulus in Florianópolis, BrazilDocument9 paginiPopulation dynamics, reproductive biology and ichthyoplankton of Cetengraulis edentulus in Florianópolis, BrazilclaudiousdecastroÎncă nu există evaluări
- Fox Feces Reveal Mammal DiversityDocument8 paginiFox Feces Reveal Mammal DiversityAntonio MangioneÎncă nu există evaluări
- Reproduction of Rough Coffee Snake and Diet of Patagonian RacerDocument2 paginiReproduction of Rough Coffee Snake and Diet of Patagonian RacerArco Iris CallejaÎncă nu există evaluări
- Water beetles in mountainous regions of southeastern BrazilDocument20 paginiWater beetles in mountainous regions of southeastern Brazilthaisa christineÎncă nu există evaluări
- Reptilia, Squamata, Amphisbaenidae, Leposternon SPP.: Distribution Extension, New State Record, and Geographic Distribution MapDocument4 paginiReptilia, Squamata, Amphisbaenidae, Leposternon SPP.: Distribution Extension, New State Record, and Geographic Distribution MapkellymafraÎncă nu există evaluări
- Artículo Original/ Original ArticleDocument9 paginiArtículo Original/ Original ArticleCami ChavezÎncă nu există evaluări
- Cunha Et AlDocument9 paginiCunha Et Alapi-3828346Încă nu există evaluări
- Santiago Alarcon& Parker2007Document11 paginiSantiago Alarcon& Parker2007Diego Santiago AlarconÎncă nu există evaluări
- RossiSantos2009 CetaceanInteractionsDocument7 paginiRossiSantos2009 CetaceanInteractionsjenn.aereaÎncă nu există evaluări
- Mastozoología Neotropical 0327-9383: Issn: Ulyses@cenpat - Edu.arDocument13 paginiMastozoología Neotropical 0327-9383: Issn: Ulyses@cenpat - Edu.arpaco jonesÎncă nu există evaluări
- Reproductive Biology of A Bromeligenous Frog Endemic To The Atlantic ForestDocument14 paginiReproductive Biology of A Bromeligenous Frog Endemic To The Atlantic ForestamandasantiagoÎncă nu există evaluări
- Ecologia Das Comunidades de Metazoários ParasitosDocument12 paginiEcologia Das Comunidades de Metazoários ParasitosRafael Souza AntunesÎncă nu există evaluări
- Alvez-Ferreira Et Al 2021 Morphological Characterization and Diversity of Tadpoles at PNEDocument11 paginiAlvez-Ferreira Et Al 2021 Morphological Characterization and Diversity of Tadpoles at PNEFausto NomuraÎncă nu există evaluări
- PopulationEcologyoftheFiddlerCrabUcaspp Inbrgy PuntaBaybayLeyteDocument14 paginiPopulationEcologyoftheFiddlerCrabUcaspp Inbrgy PuntaBaybayLeyteMelvin Earl AgdaÎncă nu există evaluări
- Vitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilDocument24 paginiVitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilEd MyersÎncă nu există evaluări
- Litter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Document12 paginiLitter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Tayssa MarquesÎncă nu există evaluări
- Reproductive Ecology of The Parthenogenetic Whiptail Cnemidophorus NativoDocument4 paginiReproductive Ecology of The Parthenogenetic Whiptail Cnemidophorus NativoRenara SantosÎncă nu există evaluări
- Elasmobranchs From Patos Lagoon Estuary: Occurrence of Catches and Population StructureDocument26 paginiElasmobranchs From Patos Lagoon Estuary: Occurrence of Catches and Population StructureThiago ChagasÎncă nu există evaluări
- Reproductive Period and Geographic Distribution of the Toad Incilius aucoinae in Golfito, Costa RicaDocument12 paginiReproductive Period and Geographic Distribution of the Toad Incilius aucoinae in Golfito, Costa RicaVanessa Usbeck LópezÎncă nu există evaluări
- Original Article / Artículo OriginalDocument13 paginiOriginal Article / Artículo OriginalYenniffer Vicente MayaÎncă nu există evaluări
- Skidds 2007Document13 paginiSkidds 2007RAKA RYANDRAÎncă nu există evaluări
- Batisteli Et Al. (2020) Breeding Biology of The Creamy-Bellied Thrush (Turdus Amaurochalinus) in Southeast BrazilDocument10 paginiBatisteli Et Al. (2020) Breeding Biology of The Creamy-Bellied Thrush (Turdus Amaurochalinus) in Southeast BrazilPaola ResendeÎncă nu există evaluări
- Bastos & Haddad, 1996 - Breeding Activity of The Neotropical Treefrog Hyla ElegansDocument7 paginiBastos & Haddad, 1996 - Breeding Activity of The Neotropical Treefrog Hyla ElegansLucas GrandinettiÎncă nu există evaluări
- Phylogeography of The Endangered Sand Dune Whiptail Lizard Glaucomastix Abaetensis (Dias, Rocha & Vrcibradic, 2002) With The Description of A New SpeciesDocument27 paginiPhylogeography of The Endangered Sand Dune Whiptail Lizard Glaucomastix Abaetensis (Dias, Rocha & Vrcibradic, 2002) With The Description of A New SpeciesIgor RosárioÎncă nu există evaluări
- Eleutherodactylus Johnstonei in An IntroducedDocument4 paginiEleutherodactylus Johnstonei in An IntroducedDavid AndresÎncă nu există evaluări
- Camargo Et Al 2004Document10 paginiCamargo Et Al 2004rini susilowatiÎncă nu există evaluări
- Seasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientDocument21 paginiSeasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientVíctor RamirezdÎncă nu există evaluări
- Callico Fortunato-G Castro Et Al - FishRes - 2017Document9 paginiCallico Fortunato-G Castro Et Al - FishRes - 2017mÎncă nu există evaluări
- 2001-Hero-et-al-Biotropica PredaçãoDocument11 pagini2001-Hero-et-al-Biotropica PredaçãoJoseane de Souza CardosoÎncă nu există evaluări
- Hyla Faber Aggressive BehaviorDocument9 paginiHyla Faber Aggressive BehaviorJoseane de Souza CardosoÎncă nu există evaluări
- Telecurso 2000 - Inglês 17Document4 paginiTelecurso 2000 - Inglês 17Ingles TCÎncă nu există evaluări
- Hyla Faber Aggressive BehaviorDocument9 paginiHyla Faber Aggressive BehaviorJoseane de Souza CardosoÎncă nu există evaluări
- A Primer For The Morphology of TadpoleDocument4 paginiA Primer For The Morphology of TadpoleJoseane de Souza CardosoÎncă nu există evaluări
- A Comparative Test of The Adaptive Plasticity Hypothesis in Anuran LarvaeDocument16 paginiA Comparative Test of The Adaptive Plasticity Hypothesis in Anuran LarvaeJoseane de Souza CardosoÎncă nu există evaluări
- Description of The Tadpole of Megaelosia Goeldii (Leptodactylidae, Hylodinae) With Natural HistorDocument6 paginiDescription of The Tadpole of Megaelosia Goeldii (Leptodactylidae, Hylodinae) With Natural HistorJoseane de Souza CardosoÎncă nu există evaluări
- Asian Torrent Frog DevelopmentDocument9 paginiAsian Torrent Frog DevelopmentJoseane de Souza CardosoÎncă nu există evaluări
- Toledo & Haddad 2005 Biology Scinax FuscomarginatusDocument9 paginiToledo & Haddad 2005 Biology Scinax FuscomarginatusJoseane de Souza CardosoÎncă nu există evaluări
- 10 Nunes e Pombal S JuncaeDocument7 pagini10 Nunes e Pombal S JuncaeJoseane de Souza CardosoÎncă nu există evaluări
- ALves-Silva & Silva - 2009 - Life in Bromeliads Reproductive Behaviour and The Monophyly of The Scinax Us Species GroupDocument14 paginiALves-Silva & Silva - 2009 - Life in Bromeliads Reproductive Behaviour and The Monophyly of The Scinax Us Species GroupJoseane de Souza CardosoÎncă nu există evaluări
- 4 - Measuring and Monitoring Biological Diversity. Standard Methods For Amphibians - Cap 2 - Mcdiarmind & Heyer - Amphibian Diversity and Natural HistoryDocument12 pagini4 - Measuring and Monitoring Biological Diversity. Standard Methods For Amphibians - Cap 2 - Mcdiarmind & Heyer - Amphibian Diversity and Natural HistoryJoseane de Souza CardosoÎncă nu există evaluări
- Herpetofauna Serrapilheira Cotia SPDocument20 paginiHerpetofauna Serrapilheira Cotia SPangstron328Încă nu există evaluări
- The Frog Scientist Discussion GuideDocument4 paginiThe Frog Scientist Discussion GuideHoughton Mifflin HarcourtÎncă nu există evaluări
- B2 TestDocument6 paginiB2 TestPhu NguyenÎncă nu există evaluări
- Amphibian Conservation Research Guide (2009) - Robert Browne.Document40 paginiAmphibian Conservation Research Guide (2009) - Robert Browne.Craig Hassapakis100% (1)
- Cut The Head Off The Frog LabDocument7 paginiCut The Head Off The Frog Labsuhabio0% (1)
- Stories For EnglishDocument36 paginiStories For Englishyannie100% (1)
- A. Multiple Choice Questions (1X10 10 Marks)Document2 paginiA. Multiple Choice Questions (1X10 10 Marks)Naresh RajuÎncă nu există evaluări
- Learning Module KSSR World of Science Technology Year 2 1st EditionDocument72 paginiLearning Module KSSR World of Science Technology Year 2 1st EditionPoospa KumaranÎncă nu există evaluări
- Freshwater EcosystemDocument22 paginiFreshwater EcosystemRitvik RitvikÎncă nu există evaluări
- Fish ListDocument12 paginiFish ListA'Jadang GfiÎncă nu există evaluări
- Sample Lab ReportDocument6 paginiSample Lab Reportvq77q9rf2tÎncă nu există evaluări
- Animals' Body Structures Help Them Adapt to HabitatsDocument3 paginiAnimals' Body Structures Help Them Adapt to Habitatsarellano lawschool100% (2)
- Nature Science4 SyllabusDocument22 paginiNature Science4 Syllabuskata perákÎncă nu există evaluări
- The Presentation Theme: A Moche Fineline DrawingDocument33 paginiThe Presentation Theme: A Moche Fineline DrawingCliffÎncă nu există evaluări
- แนวข้อสอบ O-Net วิชา ภาษาอังกฤษ มัธยมศึกษาตอนต้นDocument84 paginiแนวข้อสอบ O-Net วิชา ภาษาอังกฤษ มัธยมศึกษาตอนต้นmrlog100% (1)
- Zoo Lab Frog ActivityDocument12 paginiZoo Lab Frog ActivityAdznaira AmilussinÎncă nu există evaluări
- Practice Test 13aDocument8 paginiPractice Test 13aNhin NhanÎncă nu există evaluări
- Biodiversity (Folio)Document21 paginiBiodiversity (Folio)znha7480% (5)
- Cowichan Green Map - FULLDocument2 paginiCowichan Green Map - FULLinfo6077Încă nu există evaluări
- Frog DissectionDocument30 paginiFrog DissectionAubrey Sophia Magdasoc MisolaniaÎncă nu există evaluări
- Pineda & Halffter 2004. Species Diversity and Habitat Fragmentation: Frogs in A Tropical Montane Landscape in MexicoDocument10 paginiPineda & Halffter 2004. Species Diversity and Habitat Fragmentation: Frogs in A Tropical Montane Landscape in MexicoEduardoO_PinedaÎncă nu există evaluări
- Galactic Cannibalism CapturedDocument6 paginiGalactic Cannibalism CapturedLakshminarayan RamannaÎncă nu există evaluări
- AmphibiansDocument12 paginiAmphibianskldwujezycznaÎncă nu există evaluări
- Used To Compare and Contrast Ideas and Events - Uses Two or More Overlapping Circles To Show Similar and Different AttributesDocument10 paginiUsed To Compare and Contrast Ideas and Events - Uses Two or More Overlapping Circles To Show Similar and Different AttributesFretz AelÎncă nu există evaluări
- AMPHIBIANS - Notes, Biology GCSE - DETAILED!!Document2 paginiAMPHIBIANS - Notes, Biology GCSE - DETAILED!!Elizabeth KálusováÎncă nu există evaluări
- G4 Second-Quarter SummativeDocument28 paginiG4 Second-Quarter SummativeAllan Roy CandelariaÎncă nu există evaluări
- 6 - Experiment 2 - Physiology of The NerveDocument11 pagini6 - Experiment 2 - Physiology of The Nervepinky peachÎncă nu există evaluări
- Tetrapods + AmniotesDocument40 paginiTetrapods + Amnioteskfagg0tÎncă nu există evaluări
- The Princess and The FrogDocument4 paginiThe Princess and The Frogsimbenia_2135Încă nu există evaluări
- Frog Life Cycle - Lesson 1Document5 paginiFrog Life Cycle - Lesson 1mariamÎncă nu există evaluări
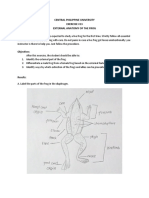
- Central Philippine University Exercise #13 External Anatomy of The FrogDocument4 paginiCentral Philippine University Exercise #13 External Anatomy of The FrogDaks ZabateÎncă nu există evaluări