S-ar putea să vă placă și
- Cold Spring Harb Perspect Biol-2010-Lee-a003236Document20 paginiCold Spring Harb Perspect Biol-2010-Lee-a003236lauranachefÎncă nu există evaluări
- Mol Cancer Ther 2006 Aziz 1335 41Document8 paginiMol Cancer Ther 2006 Aziz 1335 41Sava1988Încă nu există evaluări
- Colorectal CarcinomaDocument9 paginiColorectal CarcinomaKantona FrankÎncă nu există evaluări
- Chapitre de Livre BoutainaDocument37 paginiChapitre de Livre BoutainaBoutaina AddoumÎncă nu există evaluări
- Rare Mutations in The PIK3CA Gene Contribute To Aggressive Endometrial CancerDocument7 paginiRare Mutations in The PIK3CA Gene Contribute To Aggressive Endometrial CancerFerdina NidyasariÎncă nu există evaluări
- Módulo 23 - Licopeno - Actividad Sobre Cáncer de PróstataDocument14 paginiMódulo 23 - Licopeno - Actividad Sobre Cáncer de PróstataJany PiccoÎncă nu există evaluări
- Ni Hms 511528Document19 paginiNi Hms 511528Anonymous PKE8zOXÎncă nu există evaluări
- Epigenetic and Cancer Therapy, 2019Document12 paginiEpigenetic and Cancer Therapy, 2019AbsjeyÎncă nu există evaluări
- Zhao' 07Document11 paginiZhao' 07Javier Cerda InfanteÎncă nu există evaluări
- Molecular Mechanism of Triple Negative Breast Cancer Associated BRCA1 and The Identification of Signaling PathwaysDocument10 paginiMolecular Mechanism of Triple Negative Breast Cancer Associated BRCA1 and The Identification of Signaling PathwaysnagarajharishÎncă nu există evaluări
- Pi3k ReviewDocument14 paginiPi3k ReviewsantosdepetrisÎncă nu există evaluări
- Alterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasDocument11 paginiAlterations in The p16 /CDKN2A Tumor Suppressor Gene in GastrinomasAbdur Rachman Ba'abdullahÎncă nu există evaluări
- Azim 2016Document10 paginiAzim 2016Jocilene Dantas Torres NascimentoÎncă nu există evaluări
- High Expression of Oxidative Phosphorylation Genes Predicts Improved Survival in Squamous Cell Carcinomas of The Head and Neck and LungDocument14 paginiHigh Expression of Oxidative Phosphorylation Genes Predicts Improved Survival in Squamous Cell Carcinomas of The Head and Neck and Lungsalmaa safiraÎncă nu există evaluări
- Role of Rbm3 Gene in CancerDocument40 paginiRole of Rbm3 Gene in CancerAnkit AgarwalÎncă nu există evaluări
- Ijo 40 6 1889 PDFDocument11 paginiIjo 40 6 1889 PDFshovonÎncă nu există evaluări
- Rarres53 PDFDocument17 paginiRarres53 PDFXimenaBandaÎncă nu există evaluări
- Aberrant Proteins in Cancer RamanDocument25 paginiAberrant Proteins in Cancer RamanPetquantic -Dra.Daniela Lopes (Petquantic Saúde Quantica)Încă nu există evaluări
- YuanDocument14 paginiYuandenny juliÎncă nu există evaluări
- Anticancer Efficacy of A Difluorodiarylidenyl Piperidone (HO-3867) in Human Ovarian Cancer Cells and Tumor XenograftsDocument12 paginiAnticancer Efficacy of A Difluorodiarylidenyl Piperidone (HO-3867) in Human Ovarian Cancer Cells and Tumor XenograftsElsa Natia RindianaÎncă nu există evaluări
- 2015Glomerulonephritis-Induced Changes in Urinary and Kidney MicroRNA Profiles in RatsDocument50 pagini2015Glomerulonephritis-Induced Changes in Urinary and Kidney MicroRNA Profiles in Ratstiger KeysÎncă nu există evaluări
- This Is Your Thyroid On Drugs Targetable Mutations An 2023 Surgical PatholoDocument17 paginiThis Is Your Thyroid On Drugs Targetable Mutations An 2023 Surgical PatholorubenmacaÎncă nu există evaluări
- The Hallmarks of Cancer Review: 2000 by Cell PressDocument14 paginiThe Hallmarks of Cancer Review: 2000 by Cell PressAashiq HussainÎncă nu există evaluări
- Drosophila: Modeling Tumor Invasion and Metastasis inDocument9 paginiDrosophila: Modeling Tumor Invasion and Metastasis inFernando Lazzarotto EvaldtÎncă nu există evaluări
- NGS and Mutational Profile Analysis of Non-Small-Cell Lung Carcinoma (NSCLC)Document7 paginiNGS and Mutational Profile Analysis of Non-Small-Cell Lung Carcinoma (NSCLC)IJRASETPublicationsÎncă nu există evaluări
- Please, Some Vocabulary No Need To Change Like (Purification, SynthesisDocument6 paginiPlease, Some Vocabulary No Need To Change Like (Purification, SynthesisasdefenceÎncă nu există evaluări
- Creatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Document4 paginiCreatine Phosphokinase Levels in Oral Cancer Patients: Issn 0974-3618 (Print) 0974-360X (Online)Deepak KumarÎncă nu există evaluări
- Hallmark of CancerDocument14 paginiHallmark of CanceriinsabatiniÎncă nu există evaluări
- Erc EndocrinologyDocument11 paginiErc EndocrinologyEsti Nur EkasariÎncă nu există evaluări
- 1953 FullDocument10 pagini1953 FullboboyoyotÎncă nu există evaluări
- Chapter 1 - The Cancer GenomeDocument35 paginiChapter 1 - The Cancer GenomeCynthia LopesÎncă nu există evaluări
- Tumor ProgressionDocument7 paginiTumor ProgressionmineresearchÎncă nu există evaluări
- Reading Rnas in The Cell: From Lee, J. H., Et Al., Science, 2014, 343, 1360. Reprinted With Permission From AAASDocument3 paginiReading Rnas in The Cell: From Lee, J. H., Et Al., Science, 2014, 343, 1360. Reprinted With Permission From AAASWa RioÎncă nu există evaluări
- Animal TestingDocument15 paginiAnimal TestingMnJ9091Încă nu există evaluări
- K1-Perspectife, Blok Onk DefiDocument35 paginiK1-Perspectife, Blok Onk Defiparik2321Încă nu există evaluări
- Mutations in The EGFR Kinase Domain Mediate STAT3 Activation Via IL-6 Production in Human Lung AdenocarcinomasDocument11 paginiMutations in The EGFR Kinase Domain Mediate STAT3 Activation Via IL-6 Production in Human Lung AdenocarcinomasNilabh RanjanÎncă nu există evaluări
- Role of Matrix Metalloproteinases in Invasion and Metastasis: Biology, Diagnosis and InhibitorsDocument18 paginiRole of Matrix Metalloproteinases in Invasion and Metastasis: Biology, Diagnosis and Inhibitorsshunmu30Încă nu există evaluări
- Characterization of Soy-Based Changes in Wnt-Frizzled Signaling in Prostate CancerDocument8 paginiCharacterization of Soy-Based Changes in Wnt-Frizzled Signaling in Prostate CancerfiloshÎncă nu există evaluări
- Márcia - Epigenetica e Cancer Humano - Grupo 3Document8 paginiMárcia - Epigenetica e Cancer Humano - Grupo 3Anjas WilapanggaÎncă nu există evaluări
- Targe Ted The Rapies inDocument15 paginiTarge Ted The Rapies inAlejandro Estrada RiosÎncă nu există evaluări
- Prostate Cancer and microRNAs: New Insights Into ApoptosisDocument19 paginiProstate Cancer and microRNAs: New Insights Into ApoptosisWaldo IzyÎncă nu există evaluări
- Genetic ExpressionsDocument6 paginiGenetic ExpressionsRobertaÎncă nu există evaluări
- NIH Public Access: Author ManuscriptDocument17 paginiNIH Public Access: Author ManuscriptmarcussiÎncă nu există evaluări
- Signaling Cancer SpecialDocument4 paginiSignaling Cancer SpecialcadimogirlÎncă nu există evaluări
- Zhong 2Document6 paginiZhong 2naveedbioÎncă nu există evaluări
- p53 Enters The MicroRNA WorldDocument5 paginip53 Enters The MicroRNA WorldKenanÎncă nu există evaluări
- Targeting Signal Transduction: George Weber, Fei Shen, Tam !as I. Orb!an, Szabolcs K .Okeny, Edith OlahDocument10 paginiTargeting Signal Transduction: George Weber, Fei Shen, Tam !as I. Orb!an, Szabolcs K .Okeny, Edith Olahviralnanobio_4150420Încă nu există evaluări
- Mir-222 Overexpression Confers Cell Migratory Advantages in Hepatocellular Carcinoma Through Enhancing Akt SignalingDocument10 paginiMir-222 Overexpression Confers Cell Migratory Advantages in Hepatocellular Carcinoma Through Enhancing Akt SignalingRodrigo Costa ZeferinoÎncă nu există evaluări
- Jcem 5399Document6 paginiJcem 5399Mia DangaÎncă nu există evaluări
- The Evolving Role of The Endocannabinoid System in Gynaecological CancerDocument19 paginiThe Evolving Role of The Endocannabinoid System in Gynaecological CancernusantaramedikasolusindoÎncă nu există evaluări
- Acquired Capability: Evading ApoptosisDocument5 paginiAcquired Capability: Evading ApoptosisKaren Joy MariñasÎncă nu există evaluări
- Molecular Biomarkers of Bladder Cancer: A Mini-ReviewDocument10 paginiMolecular Biomarkers of Bladder Cancer: A Mini-ReviewaldyÎncă nu există evaluări
- 2002 DavidsonDocument5 pagini2002 DavidsonBianca PapadopolÎncă nu există evaluări
- EGFR in Meningoma 2Document5 paginiEGFR in Meningoma 2Apriyanto LatiefÎncă nu există evaluări
- Ptenless Means More: Bangyan Stiles, Matthias Groszer, Shunyou Wang, Jing Jiao, Hong WuDocument10 paginiPtenless Means More: Bangyan Stiles, Matthias Groszer, Shunyou Wang, Jing Jiao, Hong Wurocambolescas perthÎncă nu există evaluări
- Glycogen Synthase Kinase 3β Inhibition as a Therapeutic Approach in the Treatment of Endometrial CancerDocument25 paginiGlycogen Synthase Kinase 3β Inhibition as a Therapeutic Approach in the Treatment of Endometrial CancerFerdina NidyasariÎncă nu există evaluări
- Articulo 1Document6 paginiArticulo 1Fabio Andrés EnríquezÎncă nu există evaluări
- Large-Scale Screening For Somatic Mutations in Lung CancerDocument3 paginiLarge-Scale Screening For Somatic Mutations in Lung CancerAriÎncă nu există evaluări
- Xin Yan Et Al - External Qi of Yan Xin Qigong Differentially Regulates The Akt and Extracellular Signal-Regulated Kinase Pathways and Is Cytotoxic To Cancer Cells But Not To Normal CellsDocument12 paginiXin Yan Et Al - External Qi of Yan Xin Qigong Differentially Regulates The Akt and Extracellular Signal-Regulated Kinase Pathways and Is Cytotoxic To Cancer Cells But Not To Normal CellsSonyRed100% (1)
- Evolutionary Foundations For Cancer BiologyDocument16 paginiEvolutionary Foundations For Cancer BiologyDenish MikaÎncă nu există evaluări
- Conveying Misinformation: Top-Ranked Japanese Books On TobaccoDocument6 paginiConveying Misinformation: Top-Ranked Japanese Books On TobaccoDenish MikaÎncă nu există evaluări
- Book Review Of: "Clinical Aspects of Electroporation " by Stephen T Kee, Julie Gehl, Edward W LeeDocument4 paginiBook Review Of: "Clinical Aspects of Electroporation " by Stephen T Kee, Julie Gehl, Edward W LeeDenish MikaÎncă nu există evaluări
- Acacia LeucopholiaDocument6 paginiAcacia LeucopholiaDenish MikaÎncă nu există evaluări
- Tonb-Linked Outer Membrane Protein Molecular Cloning of A: Bacteroides CaccaeDocument12 paginiTonb-Linked Outer Membrane Protein Molecular Cloning of A: Bacteroides CaccaeDenish MikaÎncă nu există evaluări
- REVISION-đã Chuyển ĐổiDocument16 paginiREVISION-đã Chuyển ĐổiDrioungÎncă nu există evaluări
- The Silence-The Asbestos Industry and Early Occupational Cancer Research-A Case Study, Lilienfeld, AmJPubH-1991Document10 paginiThe Silence-The Asbestos Industry and Early Occupational Cancer Research-A Case Study, Lilienfeld, AmJPubH-1991PeopleSpeakÎncă nu există evaluări
- Akshara Kashayam (English)Document23 paginiAkshara Kashayam (English)Anthavasi100% (1)
- Budwig Diet PDFDocument4 paginiBudwig Diet PDFAlex Dcosta100% (1)
- Practice QuestionsDocument10 paginiPractice Questionsdabervid230% (1)
- Oncology Nursing: Detection and Prevention of CancerDocument14 paginiOncology Nursing: Detection and Prevention of CancerEdwin Delos Reyes AbuÎncă nu există evaluări
- 2016 EdExcel Biology Topic 5 Health Disease and MedicinesDocument40 pagini2016 EdExcel Biology Topic 5 Health Disease and MedicinesShilpa RKÎncă nu există evaluări
- Unit 5: Diagnosis Part 2: Medical History Identify UtilizeDocument4 paginiUnit 5: Diagnosis Part 2: Medical History Identify UtilizeAnonymous Wfl201YbYoÎncă nu există evaluări
- Viewpoint Synthesis Outline 1Document14 paginiViewpoint Synthesis Outline 1api-495027984Încă nu există evaluări
- Figo Hysterectomy Pelvic (Peritoneal) WashingsDocument15 paginiFigo Hysterectomy Pelvic (Peritoneal) WashingsNina Glaiza GotoÎncă nu există evaluări
- Teaching Care Plan For CancerDocument6 paginiTeaching Care Plan For CancerAinah Kristia Manzon67% (3)
- Acute Lymphoblastic LeukaemiaDocument32 paginiAcute Lymphoblastic LeukaemiaHealth Education Library for PeopleÎncă nu există evaluări
- Divine Healing Codes and How To Use Them v1.0Document34 paginiDivine Healing Codes and How To Use Them v1.0Henrii Arias83% (6)
- Marginal Zone Lymphomas: Subtypes: MALT (Extranodal), Nodal, SplenicDocument2 paginiMarginal Zone Lymphomas: Subtypes: MALT (Extranodal), Nodal, SplenicwwxxmmÎncă nu există evaluări
- Nutritional Problems in IndiaDocument87 paginiNutritional Problems in Indiagrace williamÎncă nu există evaluări
- ADULT WILMS' TUMOUR PPT - FDocument16 paginiADULT WILMS' TUMOUR PPT - FJitendra YedeÎncă nu există evaluări
- Oncology CasesDocument96 paginiOncology CasesPrutha Vaidya100% (1)
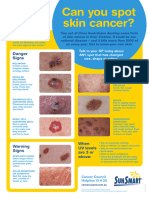
- Can You Spot Skin Cancer A3 PosterDocument1 paginăCan You Spot Skin Cancer A3 Posteralandanwar266Încă nu există evaluări
- Applications of Tomotherapy PDFDocument4 paginiApplications of Tomotherapy PDFMalu Paras LacsonÎncă nu există evaluări
- Adults Brain Tumor in Cipto Mangunkusumo General Hospital A Descriptive EpidemiologyDocument5 paginiAdults Brain Tumor in Cipto Mangunkusumo General Hospital A Descriptive EpidemiologyHarun NAÎncă nu există evaluări
- Ivermectin For CancerDocument15 paginiIvermectin For CancerSheena FriesenÎncă nu există evaluări
- Amelanotic Melanoma of The TongueDocument17 paginiAmelanotic Melanoma of The TonguedennisÎncă nu există evaluări
- Cancer QuizDocument7 paginiCancer QuizJoshua Flores Fernan100% (1)
- Wiley Protocol Consumer Newsletter DecemberDocument5 paginiWiley Protocol Consumer Newsletter DecemberWileyProtocolÎncă nu există evaluări
- PenisDocument23 paginiPenisWira DharmaÎncă nu există evaluări
- Harvard Medical School - High Impact Cancer Research Program BrochureDocument16 paginiHarvard Medical School - High Impact Cancer Research Program BrochurePUTRI NOVI SITA DEWIÎncă nu există evaluări
- Aromatherapy, Massage PDFDocument233 paginiAromatherapy, Massage PDFGyarmati LászlóÎncă nu există evaluări
- CRC and Other Colorectal Disease - PPT For Senior Medical StudentsDocument37 paginiCRC and Other Colorectal Disease - PPT For Senior Medical StudentsGiovanni HenryÎncă nu există evaluări
- Papaya Leaf Cancer CureDocument34 paginiPapaya Leaf Cancer CureFrancisca_nida75% (4)
- Colon Tumor Carcinoma WHO 2010Document3 paginiColon Tumor Carcinoma WHO 2010Ronald MoraÎncă nu există evaluări