S-ar putea să vă placă și
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoDe la EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoEvaluare: 4 din 5 stele4/5 (1)
- Calculo ChumbadoresDocument24 paginiCalculo ChumbadoresERFerrandin100% (3)
- Hyster h155ftDocument8 paginiHyster h155ftVitorMello50% (2)
- Introdução à Citometria de Fluxo: Um manual básico para iniciantesDe la EverandIntrodução à Citometria de Fluxo: Um manual básico para iniciantesÎncă nu există evaluări
- Instrumentação Industrial - PressãoDocument73 paginiInstrumentação Industrial - PressãoJohnata FrançaÎncă nu există evaluări
- Relatório Aula Prática MitoseDocument9 paginiRelatório Aula Prática MitoseRenas Pereira73% (11)
- Biologia Terceiro AnoDocument147 paginiBiologia Terceiro AnoinessantosdesouzaÎncă nu există evaluări
- Atendimento de FarmaciaDocument16 paginiAtendimento de FarmaciaJackson Da Silveira DóriaÎncă nu există evaluări
- Raiz, Caule e FolhaDocument10 paginiRaiz, Caule e FolhaFabi PaolaÎncă nu există evaluări
- Livro de Dinâmica Das Máquinas - Capitulo 1Document22 paginiLivro de Dinâmica Das Máquinas - Capitulo 1Aldair Soares0% (2)
- Radiação e decaimento radioativoDocument4 paginiRadiação e decaimento radioativoRodolfo FariasÎncă nu există evaluări
- Estudo de Caso Fachada Ventilada PDFDocument17 paginiEstudo de Caso Fachada Ventilada PDFwagsantanaÎncă nu există evaluări
- Atividade Prática - Ciências Moleculares e CelularesDocument22 paginiAtividade Prática - Ciências Moleculares e CelularesLuiz Nazareno Souza100% (1)
- RE 899-2003 - ValidaçãoDocument13 paginiRE 899-2003 - ValidaçãoGabriel BiancoÎncă nu există evaluări
- ATIVIDADE PRÁTICA SUPERVISIONADA - ConcluídaDocument3 paginiATIVIDADE PRÁTICA SUPERVISIONADA - ConcluídaAlexander CarcelenÎncă nu există evaluări
- Citometria de FluxoDocument5 paginiCitometria de FluxoGabriela SampaioÎncă nu există evaluări
- TEMA No 6. - CRECIMIENTO MICROBIANODocument18 paginiTEMA No 6. - CRECIMIENTO MICROBIANOFernanda JaldinÎncă nu există evaluări
- Citometria de fluxo na hematologia veterináriaDocument8 paginiCitometria de fluxo na hematologia veterináriaAlex Barbosa Dos SantosÎncă nu există evaluări
- Métodos diretos e indiretos para quantificação celularDocument4 paginiMétodos diretos e indiretos para quantificação celularJéssica Barbosa LimaÎncă nu există evaluări
- Relatório Completo - VirologiaDocument35 paginiRelatório Completo - VirologiapaulavonÎncă nu există evaluări
- l3Document22 paginil3anamaterialfarmaciaÎncă nu există evaluări
- Processos Fermentativos Gerais PDFDocument52 paginiProcessos Fermentativos Gerais PDFLuana LimaÎncă nu există evaluări
- RelatórioDocument8 paginiRelatóriojuanitogilsa7334Încă nu există evaluări
- Citometria PowerpointDocument20 paginiCitometria Powerpointbim380Încă nu există evaluări
- Apostila Bertho Citometria de FluxoDocument50 paginiApostila Bertho Citometria de Fluxoworkshopdengue50% (2)
- Aula 1 - IntroduçãoDocument41 paginiAula 1 - IntroduçãoLeticia RodriguesÎncă nu există evaluări
- CF CBHS - Curso 1 - 2018 - MonografiaDocument48 paginiCF CBHS - Curso 1 - 2018 - MonografiaPaula MadureuraÎncă nu există evaluări
- Técnicas de Quantificação CelularDocument20 paginiTécnicas de Quantificação CelularMarcos SilvaÎncă nu există evaluări
- Técnicas citológicas para Biologia CelularDocument4 paginiTécnicas citológicas para Biologia CelularTiago Sabença0% (1)
- Questoes 3Document9 paginiQuestoes 3LUISÎncă nu există evaluări
- Crescimento Microbiano FatoresDocument10 paginiCrescimento Microbiano FatoresGustavo SouzaÎncă nu există evaluări
- Contagem de Microrganismos em PlacasDocument4 paginiContagem de Microrganismos em PlacasDaniela Lumertz da Luz100% (2)
- Aula Prática - Cultura de CélulasDocument24 paginiAula Prática - Cultura de CélulasDiana SimõesÎncă nu există evaluări
- Manual de Automatização HematologiaDocument11 paginiManual de Automatização HematologiaGeorge Da JosinaÎncă nu există evaluări
- Atividade Citologia Histo e Embriologia 1Document21 paginiAtividade Citologia Histo e Embriologia 1Odileide Santos Batista100% (2)
- Principios Dos Contadores Automáticos - PDFDocument7 paginiPrincipios Dos Contadores Automáticos - PDFbernardo208Încă nu există evaluări
- Trabalho de Microbiologia1Document7 paginiTrabalho de Microbiologia1Paola W.sÎncă nu există evaluări
- Capítulo 4: 4.1. Estrutura E Metabolismo Da Célula 4.1.1. Estrutura Básica Da CélulaDocument37 paginiCapítulo 4: 4.1. Estrutura E Metabolismo Da Célula 4.1.1. Estrutura Básica Da CélulaKevellyn SpirandeliÎncă nu există evaluări
- Sebenta FFUL BioCel Pedro JogoDocument172 paginiSebenta FFUL BioCel Pedro JogomarianapbpbÎncă nu există evaluări
- Revisando Citologia 1 2Document5 paginiRevisando Citologia 1 2xisbrjamillyÎncă nu există evaluări
- SEMANA 02-AevoluçãodagéneticaDocument48 paginiSEMANA 02-AevoluçãodagéneticaMarcelo BrandãoÎncă nu există evaluări
- Resenha Artigo 3Document3 paginiResenha Artigo 3Drielle CeciliaÎncă nu există evaluări
- 1.2. Biologia Exercícios Resolvidos Volume 1Document48 pagini1.2. Biologia Exercícios Resolvidos Volume 1ud9011750% (2)
- Organelas - Exercícios Com GabaritoDocument10 paginiOrganelas - Exercícios Com GabaritoJuliana ScanoniÎncă nu există evaluări
- Capitulo 1 - Arthur LeskDocument66 paginiCapitulo 1 - Arthur LeskGil S. AraújoÎncă nu există evaluări
- Relatório Da Aula PráticaDocument9 paginiRelatório Da Aula PráticaMateusÎncă nu există evaluări
- HemogramaDocument12 paginiHemogramacarlafreitas30Încă nu există evaluări
- Organelas e estruturas celularesDocument10 paginiOrganelas e estruturas celularesAmanda CamargoÎncă nu există evaluări
- Relatório de BG FinalDocument7 paginiRelatório de BG Finalflara56778Încă nu există evaluări
- Análise citológica do LCRDocument10 paginiAnálise citológica do LCRLuciano Rodrigues SimoesÎncă nu există evaluări
- Lista de Exercícios - Citologia e BiotecDocument13 paginiLista de Exercícios - Citologia e BiotecMarcella GamaÎncă nu există evaluări
- Ensino Fundamental e Médio em Colégio EstadualDocument2 paginiEnsino Fundamental e Médio em Colégio EstadualGEOVANI DA SILVA LIMAÎncă nu există evaluări
- Exercicios Resolucoes Gabarito Genetica MolecularDocument32 paginiExercicios Resolucoes Gabarito Genetica MolecularRodrigo LeiteÎncă nu există evaluări
- Interpretação do hemogramaDocument11 paginiInterpretação do hemogramaRoniAuditoreRodriguesÎncă nu există evaluări
- Artigo CBEBDocument4 paginiArtigo CBEBJLRangelÎncă nu există evaluări
- Cultura de Células - MarcadoresDocument10 paginiCultura de Células - MarcadoresJoana C Bastos100% (1)
- 2d TeoriaDocument12 pagini2d TeoriaArlindo Macedo Jr.Încă nu există evaluări
- Imuno Compactado - Compressed-250 2Document48 paginiImuno Compactado - Compressed-250 2Pâmella PicançoÎncă nu există evaluări
- Metodologia para A Contagem de Cianobactérias em Células/ml. Um Novo Desafio para o Analista de LaboratórioDocument3 paginiMetodologia para A Contagem de Cianobactérias em Células/ml. Um Novo Desafio para o Analista de LaboratórioValquria LimaÎncă nu există evaluări
- Relato de ExperíênciaDocument4 paginiRelato de Experíênciaronnimendes45Încă nu există evaluări
- Citometria de Fluxo Aplicada Ao Diagnóstico Das Leucemias.Document22 paginiCitometria de Fluxo Aplicada Ao Diagnóstico Das Leucemias.Rayne Vallada Da LuzÎncă nu există evaluări
- Técnicas Básicas para Quantificar MicrorganismosDocument42 paginiTécnicas Básicas para Quantificar MicrorganismosCamila RochaÎncă nu există evaluări
- Questões Sobre A Descoberta Da CélulaDocument2 paginiQuestões Sobre A Descoberta Da CélulaMaria Cristina Lima RosaÎncă nu există evaluări
- 5 ProtDocument5 pagini5 Protpj9psvbqb9Încă nu există evaluări
- Histologia Aplicada à Investigação no CEDOCDocument5 paginiHistologia Aplicada à Investigação no CEDOCBeatriz AzevedoÎncă nu există evaluări
- Relatório Aula 6 LBBM - Quantificação de Proteínas e Separação de Aminoácidos (G4)Document23 paginiRelatório Aula 6 LBBM - Quantificação de Proteínas e Separação de Aminoácidos (G4)gabu.sivieroÎncă nu există evaluări
- As Diretrizes Da OCDE para Testes de Produtos QuímicosDocument4 paginiAs Diretrizes Da OCDE para Testes de Produtos QuímicosVladimirÎncă nu există evaluări
- MarianeFernandezBatistaDiniz TCEDocument35 paginiMarianeFernandezBatistaDiniz TCEcsryderÎncă nu există evaluări
- Tab IRS2015Document4 paginiTab IRS2015Antonio ParreiraÎncă nu există evaluări
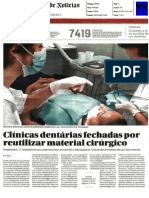
- DN ClinicasencerradasDocument3 paginiDN ClinicasencerradascsryderÎncă nu există evaluări
- Diastema InterincisivoDocument20 paginiDiastema InterincisivoNá Lopes100% (1)
- Programa Do Curso de Endodontia 2013Document7 paginiPrograma Do Curso de Endodontia 2013csryderÎncă nu există evaluări
- Controvérsias Na Anticoagulação Oral-Continuar Ou Interromper Os Anticoagulantes Orais Durante A Intervenção EstomatológicaDocument14 paginiControvérsias Na Anticoagulação Oral-Continuar Ou Interromper Os Anticoagulantes Orais Durante A Intervenção EstomatológicacsryderÎncă nu există evaluări
- Folheto informativo sobre Prednisolona LabesfalDocument7 paginiFolheto informativo sobre Prednisolona LabesfalcsryderÎncă nu există evaluări
- Diastema InterincisivoDocument20 paginiDiastema InterincisivoNá Lopes100% (1)
- Desordens Musculoesqueléticas em Cirurgiões-DentistasDocument16 paginiDesordens Musculoesqueléticas em Cirurgiões-DentistascsryderÎncă nu există evaluări
- Contactos Dentários em Pacientes Com Desordens TemporomandibularesDocument14 paginiContactos Dentários em Pacientes Com Desordens TemporomandibularescsryderÎncă nu există evaluări
- Aspectos clínicos e histológicos da cárie aguda x crônicaDocument5 paginiAspectos clínicos e histológicos da cárie aguda x crônicacsryder100% (1)
- Avaliação Da Inclinação Dentária No Tratamento Compensatório Do Padrão II Com Tomografia ComputadorizadaDocument12 paginiAvaliação Da Inclinação Dentária No Tratamento Compensatório Do Padrão II Com Tomografia ComputadorizadacsryderÎncă nu există evaluări
- 457 6890 4 PBDocument9 pagini457 6890 4 PBOtavio HenriqueÎncă nu există evaluări
- Ansiedade e Fobia Dentária - Avaliação Psicométrica Num Estudo TransversalDocument10 paginiAnsiedade e Fobia Dentária - Avaliação Psicométrica Num Estudo TransversalcsryderÎncă nu există evaluări
- Análise Crítica Dos Fatores Que Influenciam A Precisão de Moldagens Com ElastômerosDocument5 paginiAnálise Crítica Dos Fatores Que Influenciam A Precisão de Moldagens Com ElastômeroscsryderÎncă nu există evaluări
- Contactos Dentários em Pacientes Com Desordens TemporomandibularesDocument14 paginiContactos Dentários em Pacientes Com Desordens TemporomandibularescsryderÎncă nu există evaluări
- Remoção de Metais Pesados de Águas Residuais e Síntese de Nanomateriais Fotocatalíticos2Document43 paginiRemoção de Metais Pesados de Águas Residuais e Síntese de Nanomateriais Fotocatalíticos2JacksonÎncă nu există evaluări
- Escoamentos em Canais AbertosDocument637 paginiEscoamentos em Canais Abertosgabi_si2Încă nu există evaluări
- Correntes MarinhasDocument149 paginiCorrentes MarinhasAlessandro NetoÎncă nu există evaluări
- 02 Terminologia Final2Document18 pagini02 Terminologia Final2Gustavo DaviÎncă nu există evaluări
- Evolução dos Modelos AtômicosDocument18 paginiEvolução dos Modelos AtômicosThiago SantosÎncă nu există evaluări
- Fusíveis e disjuntores: funções e medidas de proteção em circuitos elétricosDocument467 paginiFusíveis e disjuntores: funções e medidas de proteção em circuitos elétricoshanameel souza100% (2)
- 2-Teste Escrito - (Grupo)Document6 pagini2-Teste Escrito - (Grupo)ARF07Încă nu există evaluări
- As Rochas São Agregados Sólidos Compostos Por Um Ou Mais MineraisDocument3 paginiAs Rochas São Agregados Sólidos Compostos Por Um Ou Mais MineraisMatheus Ribeiro OliveiraÎncă nu există evaluări
- Atividae Avaliativa 3anoDocument1 paginăAtividae Avaliativa 3anoDenise NascimentoÎncă nu există evaluări
- PlutônioDocument5 paginiPlutôniosuperpericles1Încă nu există evaluări
- Aula Bomb AsDocument116 paginiAula Bomb AsRaquel AraújoÎncă nu există evaluări
- 3340F401TDocument2 pagini3340F401TLeandro Guilherme de FariaÎncă nu există evaluări
- Tubulacoes Acessorios ValvulasDocument55 paginiTubulacoes Acessorios ValvulasMah_iseÎncă nu există evaluări
- QuestãoDocument46 paginiQuestãoSuzana MacedoÎncă nu există evaluări
- Atividades EstáticaDocument6 paginiAtividades EstáticaSilvio EduardoÎncă nu există evaluări
- 21 Ultra SomDocument9 pagini21 Ultra SomPriscila PereiraÎncă nu există evaluări
- Soldagem Oxi AcetilenoDocument3 paginiSoldagem Oxi AcetilenoFelipe MedeirosÎncă nu există evaluări
- Bibliografia SugeridaDocument3 paginiBibliografia SugeridaChico PedrosaÎncă nu există evaluări
- Roteiro de Aula PráticaDocument7 paginiRoteiro de Aula PráticaFabio WuÎncă nu există evaluări
- Lista de EPI's Homologados LHDocument46 paginiLista de EPI's Homologados LHigor AndradeÎncă nu există evaluări
- teste_modeloDocument2 paginiteste_modeloMariana MartinsÎncă nu există evaluări