S-ar putea să vă placă și
- Introduction To Molecular BiologyDocument82 paginiIntroduction To Molecular Biologyfarzaana100% (2)
- Barker CH Naish D - Where On Earth Dinosaurs and Other Prehistoric Life The Amazing History of Earth 39 S Most Incredible ADocument162 paginiBarker CH Naish D - Where On Earth Dinosaurs and Other Prehistoric Life The Amazing History of Earth 39 S Most Incredible ARumen Deshkov100% (1)
- Molecular Biology and Biotechnology: PrimerDocument48 paginiMolecular Biology and Biotechnology: PrimerQeukling143Încă nu există evaluări
- Cell and Molecular Biology For Environmental EngineersDocument44 paginiCell and Molecular Biology For Environmental EngineersCharleneKronstedtÎncă nu există evaluări
- CH 6 A Tour of The CellDocument7 paginiCH 6 A Tour of The Cellwil7verÎncă nu există evaluări
- 4 International Code On Zoological NomenclatureDocument57 pagini4 International Code On Zoological NomenclaturemuralitharangisÎncă nu există evaluări
- Protein FoldingDocument42 paginiProtein FoldingTerri PerryÎncă nu există evaluări
- Child Development An Introduction Santrock 14th Edition Test BankDocument60 paginiChild Development An Introduction Santrock 14th Edition Test BankWilliam Williams100% (36)
- Bacterial PlasmidsDocument9 paginiBacterial PlasmidsspanishinqÎncă nu există evaluări
- Yin Yang PrimerDocument53 paginiYin Yang PrimerAnonymousFarmer100% (6)
- Nucleic Acid MetabolismDocument10 paginiNucleic Acid MetabolismIvy AggabaoÎncă nu există evaluări
- Activity 7 Isolation of Bacteria Into Pure CultureDocument43 paginiActivity 7 Isolation of Bacteria Into Pure CultureglorigreizÎncă nu există evaluări
- Final Molecular BiologyDocument8 paginiFinal Molecular Biologyrachyb7100% (1)
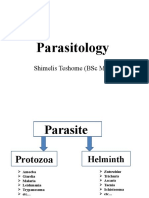
- ParasitologyDocument107 paginiParasitologyShimelis Teshome AyalnehÎncă nu există evaluări
- A Compilation of Philippine Environmental LawsDocument39 paginiA Compilation of Philippine Environmental LawsRussel Sirot100% (3)
- MCB 11 ST Q1Document25 paginiMCB 11 ST Q1Vic DBÎncă nu există evaluări
- Questions Bank For Molecular Biology Answer Sheet PDFDocument50 paginiQuestions Bank For Molecular Biology Answer Sheet PDFPranjali PriyaÎncă nu există evaluări
- Bergey S Manual of D 1957 AmerDocument1.130 paginiBergey S Manual of D 1957 AmerAbraham Calderón100% (6)
- Worksheet On Bacterial GrowthDocument5 paginiWorksheet On Bacterial GrowthBlessy MartinÎncă nu există evaluări
- Culture Media & Culture MethodsDocument60 paginiCulture Media & Culture MethodsGretchen Quimno Ayuson100% (2)
- Vectors: A Survey of Molecular Cloning Vectors and Their UsesDe la EverandVectors: A Survey of Molecular Cloning Vectors and Their UsesRaymond L. RodriguezÎncă nu există evaluări
- Histology SyllabusDocument32 paginiHistology SyllabusAdrianAddieNovioDeJesusÎncă nu există evaluări
- Exercise 8aDocument6 paginiExercise 8aKobe Conrad Abellera100% (1)
- Bio Lab 2 RespirationDocument7 paginiBio Lab 2 RespirationDominic100% (1)
- Cell Structure and Function by MicrospectrofluorometryDe la EverandCell Structure and Function by MicrospectrofluorometryÎncă nu există evaluări
- Test Bank For Microbiology A Systems Approach 3rd Edition by CowanDocument25 paginiTest Bank For Microbiology A Systems Approach 3rd Edition by CowanJoseph Williams100% (1)
- 1B. Culture MethodsDocument18 pagini1B. Culture MethodsSalman MajidÎncă nu există evaluări
- Jib 221 Microbiology: The Main Themes of MicrobiologyDocument47 paginiJib 221 Microbiology: The Main Themes of MicrobiologyNazzÎncă nu există evaluări
- Indian Constituition 1 WordsDocument9 paginiIndian Constituition 1 WordsChinnasamy100% (1)
- Nmat Msa ConceptsDocument2 paginiNmat Msa ConceptsGeovin Dexter UyÎncă nu există evaluări
- BacteriaDocument50 paginiBacteriaDev DasÎncă nu există evaluări
- Microbial Products of Potential Importance - Product / Activity Examples ProductsDocument10 paginiMicrobial Products of Potential Importance - Product / Activity Examples ProductsShveta Jaishankar Iyer100% (4)
- Microbiology ReviewerDocument16 paginiMicrobiology ReviewerAnna CastroÎncă nu există evaluări
- Early Beginning of Medical Technology: Vivian HerrickDocument13 paginiEarly Beginning of Medical Technology: Vivian HerrickWho KnowsÎncă nu există evaluări
- Recognizining Individuals of Plant PopulationDocument11 paginiRecognizining Individuals of Plant PopulationFatima GraceÎncă nu există evaluări
- Botany Chapter 1 Notes PDFDocument2 paginiBotany Chapter 1 Notes PDFSUDHIR MISHRAÎncă nu există evaluări
- Separation Techniques For ProteinsDocument2 paginiSeparation Techniques For ProteinsRobert Dominic GonzalesÎncă nu există evaluări
- KUBYDocument5 paginiKUBYCristina CisnerosÎncă nu există evaluări
- Environmental BiotechnologyDocument13 paginiEnvironmental BiotechnologydgkulkarniÎncă nu există evaluări
- Selected Topics in the History of Biochemistry. Personal Recollections. Part IIIDe la EverandSelected Topics in the History of Biochemistry. Personal Recollections. Part IIIEvaluare: 1 din 5 stele1/5 (1)
- Mutiplexpcr Primer DesignDocument11 paginiMutiplexpcr Primer DesignAnn Irene DomnicÎncă nu există evaluări
- Animal Cell and Tissue Culture ApplicationsDocument2 paginiAnimal Cell and Tissue Culture ApplicationsbhagyashreesangramÎncă nu există evaluări
- Bio MajorDocument4 paginiBio MajorAnna LiÎncă nu există evaluări
- GBPR Principles of Biotechnology PDFDocument152 paginiGBPR Principles of Biotechnology PDFSabesan TÎncă nu există evaluări
- Ubiquity of MicroorganismsDocument16 paginiUbiquity of MicroorganismsNichols Amy TarunÎncă nu există evaluări
- Micro Assign FinalDocument4 paginiMicro Assign FinalArizaldo Enriquez Castro100% (1)
- NMAT BiologyDocument6 paginiNMAT BiologyJyl Yan SelasorÎncă nu există evaluări
- Nmat - Final ExamDocument7 paginiNmat - Final ExamReyn BinanLagunaÎncă nu există evaluări
- Endosymbiotic TheoryDocument4 paginiEndosymbiotic TheoryJoshi MaitreyeeÎncă nu există evaluări
- The Pyridine Nucleotide CoenzymesDe la EverandThe Pyridine Nucleotide CoenzymesJohannes EverseÎncă nu există evaluări
- Phylogenetic Tree Creation Morphological and Molecular Methods For 07-JohnsonDocument35 paginiPhylogenetic Tree Creation Morphological and Molecular Methods For 07-JohnsonCHRISTEROP100% (2)
- CH 10Document13 paginiCH 10patilamardip0078122Încă nu există evaluări
- BE Biotechnology 2015-16Document101 paginiBE Biotechnology 2015-16Sonaal GuptaÎncă nu există evaluări
- Differential Diagnosis of Malaria ParasiteDocument101 paginiDifferential Diagnosis of Malaria ParasiteSohaib Omer Salih100% (1)
- Examn of Molecular BiologyDocument11 paginiExamn of Molecular BiologycegfracÎncă nu există evaluări
- Molecular Cell Biology 1 Basics of Molecular BiologyDocument80 paginiMolecular Cell Biology 1 Basics of Molecular BiologyIana Mira100% (2)
- History of Medical TechnologyDocument19 paginiHistory of Medical TechnologyPam FajardoÎncă nu există evaluări
- Agarose Gel ElectrophoresisDocument5 paginiAgarose Gel ElectrophoresisAyioKun100% (3)
- CH 6Document7 paginiCH 6fotero100% (1)
- M SC Microbial BiotechnologyDocument31 paginiM SC Microbial BiotechnologyChetan MohanÎncă nu există evaluări
- MeiosisDe la EverandMeiosisPeter MoensEvaluare: 5 din 5 stele5/5 (1)
- 1.3 Further ReadingDocument3 pagini1.3 Further ReadingAntonio M ChavezÎncă nu există evaluări
- History of BiochemistryDocument3 paginiHistory of BiochemistryAmisha ChakrabortyÎncă nu există evaluări
- Biochimica Et Biophysica Acta: Jasvinder Singh Bhatti, Gurjit Kaur Bhatti, P. Hemachandra ReddyDocument12 paginiBiochimica Et Biophysica Acta: Jasvinder Singh Bhatti, Gurjit Kaur Bhatti, P. Hemachandra ReddyBi AnhÎncă nu există evaluări
- University of Toronto Ontario Biology Competition: 1995 Examination Answer KeyDocument1 paginăUniversity of Toronto Ontario Biology Competition: 1995 Examination Answer KeyBi AnhÎncă nu există evaluări
- VGEC: Teacher/Student Notes: Southern Blot ProtocolDocument2 paginiVGEC: Teacher/Student Notes: Southern Blot ProtocolBi AnhÎncă nu există evaluări
- Pset 2015Document1 paginăPset 2015Bi AnhÎncă nu există evaluări
- BS2009 Practicals 2008Document4 paginiBS2009 Practicals 2008Bi AnhÎncă nu există evaluări
- Figure 7.9 Regulation of The Cell CycleDocument1 paginăFigure 7.9 Regulation of The Cell CycleBi AnhÎncă nu există evaluări
- WWD 7.1 Discovery of A Second MessengerDocument2 paginiWWD 7.1 Discovery of A Second MessengerBi AnhÎncă nu există evaluări
- E00281 FullDocument13 paginiE00281 FullBi AnhÎncă nu există evaluări
- BIOL207 Open GeneticsDocument182 paginiBIOL207 Open GeneticsBi Anh100% (1)
- WWD 7.1 Discovery of A Second MessengerDocument2 paginiWWD 7.1 Discovery of A Second MessengerBi AnhÎncă nu există evaluări
- Bio 306 General Physiology-2Document143 paginiBio 306 General Physiology-2Bi AnhÎncă nu există evaluări
- How Cells Coordinate Growth and Division ReviewDocument14 paginiHow Cells Coordinate Growth and Division ReviewBi AnhÎncă nu există evaluări
- Exam 3 2008Document9 paginiExam 3 2008Bi AnhÎncă nu există evaluări
- Theory and Measurement of Bacterial GrowthDocument11 paginiTheory and Measurement of Bacterial GrowthBi Anh100% (1)
- Summer PlaceSummer Placements List Summer 2013ments List Summer 2013Document21 paginiSummer PlaceSummer Placements List Summer 2013ments List Summer 2013Bi AnhÎncă nu există evaluări
- UoN UnderGrad StudySkills 2011Document184 paginiUoN UnderGrad StudySkills 2011Bi AnhÎncă nu există evaluări
- Section CDocument1 paginăSection CBi AnhÎncă nu există evaluări
- Article On Gilead Sciences and Its Founder and CEO Dr. Michael L. RiordanDocument7 paginiArticle On Gilead Sciences and Its Founder and CEO Dr. Michael L. RiordanDorseyMarkup100% (34)
- Phylogeny and EvolutionDocument12 paginiPhylogeny and EvolutionKathryn Palmerton0% (1)
- Mukesh BiodataDocument11 paginiMukesh Biodatashrey_jindal33% (12)
- University of Karachi: World Class Institution of Higher Education and ResearchDocument2 paginiUniversity of Karachi: World Class Institution of Higher Education and ResearchfarooqusmanÎncă nu există evaluări
- Introduction To Heterocyclic Chemistry: 1.1 CoverageDocument17 paginiIntroduction To Heterocyclic Chemistry: 1.1 CoverageFlavio SantosÎncă nu există evaluări
- Production and Marketing Systems of Sheep and Goats in Alaba, Southern EthiopiaDocument174 paginiProduction and Marketing Systems of Sheep and Goats in Alaba, Southern EthiopiaYegnanewÎncă nu există evaluări
- Westren and Northern BlottingDocument13 paginiWestren and Northern BlottingGanesh V GaonkarÎncă nu există evaluări
- Barrett's EsophagusDocument10 paginiBarrett's EsophagusaryadroettninguÎncă nu există evaluări
- Cross-Cultural Differences in The Effects of Abstract and Concrete Thinking On Imagery Generation and Ad Persuasion PDFDocument35 paginiCross-Cultural Differences in The Effects of Abstract and Concrete Thinking On Imagery Generation and Ad Persuasion PDFViolaQianÎncă nu există evaluări
- Pro-Bio Beads: Solgar Vitamin and HerbDocument2 paginiPro-Bio Beads: Solgar Vitamin and HerbM Usman KhanÎncă nu există evaluări
- 2022 - 1 Pop Final Exam Timetable: Day Date 8am 11am 2pm Tuesday, 19th April 2022Document11 pagini2022 - 1 Pop Final Exam Timetable: Day Date 8am 11am 2pm Tuesday, 19th April 2022okoro collinsÎncă nu există evaluări
- Jurnal KelDocument3 paginiJurnal Kelandre erlisÎncă nu există evaluări
- Lecture 2 - Zoology - An Ecological PerspectiveDocument23 paginiLecture 2 - Zoology - An Ecological PerspectiveAbegailÎncă nu există evaluări
- Cellular DNA Polymerases: Teresa S.-F. WangDocument33 paginiCellular DNA Polymerases: Teresa S.-F. WangSuryajeeva SuresÎncă nu există evaluări
- Herpes Simplex Virus: Herpesviridae FamilyDocument9 paginiHerpes Simplex Virus: Herpesviridae FamilyANNEÎncă nu există evaluări
- Lesson 2Document3 paginiLesson 2api-483115499Încă nu există evaluări
- Lecture 10 - Extension Mendelian Genetics 2013Document35 paginiLecture 10 - Extension Mendelian Genetics 2013Carina JLÎncă nu există evaluări
- Photosynthesis - WikipediaDocument22 paginiPhotosynthesis - Wikipediaمحمد جانÎncă nu există evaluări
- Preview Guidelines For Vegetation Management AASHTODocument13 paginiPreview Guidelines For Vegetation Management AASHTOBárbara CastroÎncă nu există evaluări
- Learning Activity Sheet: Agri-Fishery Arts 11 Agricultural Crops Production NC IIDocument12 paginiLearning Activity Sheet: Agri-Fishery Arts 11 Agricultural Crops Production NC IIKM Nicolas AgcaoiliÎncă nu există evaluări
- PCR Master Mix Protocol PDFDocument2 paginiPCR Master Mix Protocol PDFCHIRANJEEVIÎncă nu există evaluări
- The Physic Body MechanicsDocument8 paginiThe Physic Body MechanicsNur RasyiqahÎncă nu există evaluări
- Narrative Text The Lion and The MouseDocument5 paginiNarrative Text The Lion and The MouseTidak Ingin Dikenal33% (3)
- Selander 1991 - On The Nomenclature and Classification of The MeloidaeDocument31 paginiSelander 1991 - On The Nomenclature and Classification of The MeloidaecristianÎncă nu există evaluări