S-ar putea să vă placă și
- Knights Templar: Medieval Christian military orderDocument16 paginiKnights Templar: Medieval Christian military orderBearer2008Încă nu există evaluări
- Santiago de CompostelaDocument13 paginiSantiago de CompostelaBearer2008Încă nu există evaluări
- #1 - The Celestial Battle: Nibiru Meets Solaris and Tiamat (Proto Earth)Document138 pagini#1 - The Celestial Battle: Nibiru Meets Solaris and Tiamat (Proto Earth)Bearer2008Încă nu există evaluări
- 2008 Hungarian Grand PrixDocument4 pagini2008 Hungarian Grand PrixBearer2008Încă nu există evaluări
- Hedley VerityDocument4 paginiHedley VerityBearer2008Încă nu există evaluări
- Maria I of PortugalDocument11 paginiMaria I of PortugalBearer2008Încă nu există evaluări
- List of Portuguese MonarchsDocument8 paginiList of Portuguese MonarchsBearer2008Încă nu există evaluări
- Order of AvizDocument5 paginiOrder of AvizBearer2008Încă nu există evaluări
- Manuel II of PortugalDocument18 paginiManuel II of PortugalBearer2008Încă nu există evaluări
- House of BourbonDocument3 paginiHouse of BourbonBearer2008Încă nu există evaluări
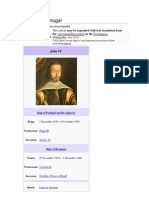
- John IV of PortugalDocument6 paginiJohn IV of PortugalBearer2008Încă nu există evaluări
- Molecular GeneticsDocument5 paginiMolecular GeneticsBearer2008Încă nu există evaluări
- Afonso VI of PortugalDocument8 paginiAfonso VI of PortugalBearer2008Încă nu există evaluări
- Prince of BrazilDocument1 paginăPrince of BrazilBearer2008Încă nu există evaluări
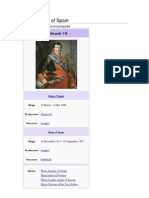
- Ferdinand VII of SpainDocument8 paginiFerdinand VII of SpainBearer2008Încă nu există evaluări
- Order of CalatravaDocument8 paginiOrder of CalatravaBearer2008Încă nu există evaluări
- ST PaulDocument7 paginiST PaulBearer2008Încă nu există evaluări
- Yersinia PestisDocument10 paginiYersinia PestisBearer2008Încă nu există evaluări
- Great Fire of LondonDocument4 paginiGreat Fire of LondonBearer2008Încă nu există evaluări
- PostmanDocument17 paginiPostmanBearer2008Încă nu există evaluări
- Yorkshire Evening PostDocument7 paginiYorkshire Evening PostBearer2008Încă nu există evaluări
- Bubonic Plague: Deadly Disease Caused by Yersinia Pestis BacteriaDocument9 paginiBubonic Plague: Deadly Disease Caused by Yersinia Pestis BacteriaBearer2008Încă nu există evaluări
- Linnean Tercentenary MedalDocument2 paginiLinnean Tercentenary MedalBearer2008Încă nu există evaluări
- Linnean Society of London: Promoting Taxonomy and BiodiversityDocument2 paginiLinnean Society of London: Promoting Taxonomy and BiodiversityBearer2008Încă nu există evaluări
- David William MitchellDocument3 paginiDavid William MitchellBearer2008Încă nu există evaluări
- George Robert GrayDocument3 paginiGeorge Robert GrayBearer2008Încă nu există evaluări
- John Edward GrayDocument5 paginiJohn Edward GrayBearer2008Încă nu există evaluări
- CoccyxDocument5 paginiCoccyxBearer2008Încă nu există evaluări
- Tort Rici DaeDocument4 paginiTort Rici DaeBearer2008Încă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- APTARE IT Analytics: Presenter NameDocument16 paginiAPTARE IT Analytics: Presenter NameCCIE DetectÎncă nu există evaluări
- Chapter 04Document3 paginiChapter 04gebreÎncă nu există evaluări
- Differentiation: Vehicle Network SolutionsDocument1 paginăDifferentiation: Vehicle Network SolutionsДрагиша Небитни ТрифуновићÎncă nu există evaluări
- Research PaperDocument15 paginiResearch PapershrirangÎncă nu există evaluări
- Strategy GlossaryDocument15 paginiStrategy GlossaryMahmoud SaeedÎncă nu există evaluări
- C++ Programmierung (Benjamin Buch, Wikibooks - Org)Document257 paginiC++ Programmierung (Benjamin Buch, Wikibooks - Org)stefano rossiÎncă nu există evaluări
- Control Systems Engineering by S C Goyal U A Bakshi PDFDocument3 paginiControl Systems Engineering by S C Goyal U A Bakshi PDFShubham SinghÎncă nu există evaluări
- Service: Audi A6 1998Document256 paginiService: Audi A6 1998Kovács EndreÎncă nu există evaluări
- Amex Case StudyDocument12 paginiAmex Case StudyNitesh JainÎncă nu există evaluări
- Case Study - Help DocumentDocument2 paginiCase Study - Help DocumentRahÎncă nu există evaluări
- Fi 7160Document2 paginiFi 7160maxis2022Încă nu există evaluări
- 6a. ICMR STSDocument15 pagini6a. ICMR STSVishnu Praba AÎncă nu există evaluări
- Case Study, g6Document62 paginiCase Study, g6julie pearl peliyoÎncă nu există evaluări
- Past Paper Booklet - QPDocument506 paginiPast Paper Booklet - QPMukeshÎncă nu există evaluări
- 1 s2.0 S0959652619316804 MainDocument11 pagini1 s2.0 S0959652619316804 MainEmma RouyreÎncă nu există evaluări
- Insert BondingDocument14 paginiInsert BondingHelpful HandÎncă nu există evaluări
- Biotechnology Eligibility Test (BET) For DBT-JRF Award (2010-11)Document20 paginiBiotechnology Eligibility Test (BET) For DBT-JRF Award (2010-11)Nandakumar HaorongbamÎncă nu există evaluări
- SIO 12 Syllabus 17Document3 paginiSIO 12 Syllabus 17Paul RobaiaÎncă nu există evaluări
- Basics of Duct DesignDocument2 paginiBasics of Duct DesignRiza BahrullohÎncă nu există evaluări
- Destroyed Inventory Deduction ProceduresDocument7 paginiDestroyed Inventory Deduction ProceduresCliff DaquioagÎncă nu există evaluări
- Digestive System Song by MR ParrDocument2 paginiDigestive System Song by MR ParrRanulfo MayolÎncă nu există evaluări
- Outstanding 12m Bus DrivelineDocument2 paginiOutstanding 12m Bus DrivelineArshad ShaikhÎncă nu există evaluări
- COP2251 Syllabus - Ellis 0525Document9 paginiCOP2251 Syllabus - Ellis 0525Satish PrajapatiÎncă nu există evaluări
- CAM TOOL Solidworks PDFDocument6 paginiCAM TOOL Solidworks PDFHussein ZeinÎncă nu există evaluări
- Monitoring Tool in ScienceDocument10 paginiMonitoring Tool in ScienceCatherine RenanteÎncă nu există evaluări
- O - 6 Series Mill Operation Manual-ENDocument119 paginiO - 6 Series Mill Operation Manual-ENLeonardo OlivaresÎncă nu există evaluări
- PCG Master Consultancy Services AgreementDocument12 paginiPCG Master Consultancy Services Agreementawscobie100% (1)
- The Collected Letters of Flann O'BrienDocument640 paginiThe Collected Letters of Flann O'BrienSean MorrisÎncă nu există evaluări
- IBM Systems Journal PerspectivesDocument24 paginiIBM Systems Journal PerspectivesSmitha MathewÎncă nu există evaluări
- Minimum Fees To Be Taken by CADocument8 paginiMinimum Fees To Be Taken by CACA Sanjay BhatiaÎncă nu există evaluări