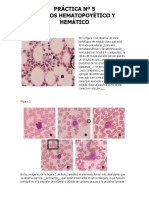
S-ar putea să vă placă și
- Fasc 7 SegmentaciónDocument31 paginiFasc 7 SegmentaciónJavier OsorioÎncă nu există evaluări
- Practica 6 Adi-Hema09Document4 paginiPractica 6 Adi-Hema09capire1111028Încă nu există evaluări
- Embriologia Marquez01Document29 paginiEmbriologia Marquez01Gerardo Gonzalez GuadarramaÎncă nu există evaluări
- Práctica 5. Tejido Hemático y Hematopoyético - EjerciciosDocument2 paginiPráctica 5. Tejido Hemático y Hematopoyético - EjerciciosDaav L. GuarneriÎncă nu există evaluări
- Programas y PracticasDocument65 paginiProgramas y PracticasITZAMARAZAVALA100% (1)
- Ovogenesis PDFDocument27 paginiOvogenesis PDFEstefani MarroquinÎncă nu există evaluări
- El Sistema Nervioso Desde La ConcepcionDocument53 paginiEl Sistema Nervioso Desde La ConcepcionALEJANDRO OSTOIC ROZZI100% (1)
- Grácil Cuneado funciones núcleosDocument10 paginiGrácil Cuneado funciones núcleosAtilio LópezÎncă nu există evaluări
- Seminario 2 Fisiologia 1Document43 paginiSeminario 2 Fisiologia 1Allison GonzalezÎncă nu există evaluări
- Fijas Cardiologia RM 2019 IntermedioDocument3 paginiFijas Cardiologia RM 2019 IntermedioThalía Muñoz RivasplataÎncă nu există evaluări
- PRÁCTICA No. 5 TALLO CEREBRAL Dr. Luis Arturo Juárez I. Marzo 2012Document10 paginiPRÁCTICA No. 5 TALLO CEREBRAL Dr. Luis Arturo Juárez I. Marzo 2012Julio PerezÎncă nu există evaluări
- EsplenosisDocument34 paginiEsplenosisMarcelo AntonioÎncă nu există evaluări
- Fisio Practica 1Document3 paginiFisio Practica 1Gerardo HernandezÎncă nu există evaluări
- Sistema cardiovascularDocument13 paginiSistema cardiovascularennys perezÎncă nu există evaluări
- Practica de Histologia - Tejido HematopoyeticoDocument4 paginiPractica de Histologia - Tejido HematopoyeticoChristian Escamilla100% (1)
- Practica Sistema EndócrinoDocument4 paginiPractica Sistema EndócrinoLili MïrandaÎncă nu există evaluări
- Embriologia Del Tronco EncefálicoDocument5 paginiEmbriologia Del Tronco EncefálicoMARIA AZUCENA CRUZADO TOLEDOÎncă nu există evaluări
- Fenomenos Mecanicos Del CorazonDocument2 paginiFenomenos Mecanicos Del CorazonGrecia GarayÎncă nu există evaluări
- El Conjunto de Órganos Nerviosos Protegidos Por El Cráneo Constituyen ElDocument6 paginiEl Conjunto de Órganos Nerviosos Protegidos Por El Cráneo Constituyen ElCristian Valbuena100% (1)
- La corriente If y su papel en la actividad de marcapasos cardiacoDocument2 paginiLa corriente If y su papel en la actividad de marcapasos cardiacoKarla HdzÎncă nu există evaluări
- EpitelialDocument18 paginiEpitelialmichaelÎncă nu există evaluări
- Rigidez por descerebraciónDocument3 paginiRigidez por descerebraciónAndrea Lucía Rodríguez100% (1)
- Cuestionario Sobre La Historia de La HistologíaDocument2 paginiCuestionario Sobre La Historia de La HistologíaGeomaraÎncă nu există evaluări
- Practica Disección de Ojo 02Document8 paginiPractica Disección de Ojo 02Laura Itzel Peñaloza GranadosÎncă nu există evaluări
- Practica #8Document23 paginiPractica #8Alondra RodriguezÎncă nu există evaluări
- Resolución de Problemas - Cap - 13Document2 paginiResolución de Problemas - Cap - 13fabiana javivaÎncă nu există evaluări
- Difusion y Medio Ambiente InternoDocument25 paginiDifusion y Medio Ambiente InternoKK-3Încă nu există evaluări
- Morfo 4 Examen FinalDocument6 paginiMorfo 4 Examen FinalwendyÎncă nu există evaluări
- TejidoOs-HistologiaDocument12 paginiTejidoOs-HistologiaBetiana VieraÎncă nu există evaluări
- Embriologia PreguntasDocument208 paginiEmbriologia PreguntasCristhian AndréÎncă nu există evaluări
- Casos Clinicos Aparato CardiovascularDocument5 paginiCasos Clinicos Aparato CardiovascularCarlos TorresÎncă nu există evaluări
- Investigues de Micro Arreglado 1Document6 paginiInvestigues de Micro Arreglado 1Albert Diaz100% (8)
- Excitación y Contracción Del Músculo Liso.Document45 paginiExcitación y Contracción Del Músculo Liso.Ambar Peña MejiaÎncă nu există evaluări
- Presentación Tumores Cerebrales. Ke y MaDocument30 paginiPresentación Tumores Cerebrales. Ke y Maluismeilyn0% (1)
- Cuestionario Receptores JJGCHDocument7 paginiCuestionario Receptores JJGCHXime Moran100% (1)
- Infografía Sobre MeiosisDocument5 paginiInfografía Sobre Meiosismateo velezÎncă nu există evaluări
- Embriologia 13Document7 paginiEmbriologia 13Duvan Zavaleta BazanÎncă nu există evaluări
- PP T 0000033Document12 paginiPP T 0000033sylviusÎncă nu există evaluări
- Guía Nº3 Tejido Epitelial GlandularDocument11 paginiGuía Nº3 Tejido Epitelial GlandularMael TamburiÎncă nu există evaluări
- Resumen HipófisisDocument8 paginiResumen HipófisisNoelia MontoyaÎncă nu există evaluări
- Trabajo Sistema de ConduccionDocument3 paginiTrabajo Sistema de Conduccionbrigithe grimaldosÎncă nu există evaluări
- El Sistema GlimfaticoDocument15 paginiEl Sistema GlimfaticoAlumnos Medicina100% (2)
- Glándulas Tiroides y ParatiroidesDocument38 paginiGlándulas Tiroides y ParatiroidesMiguel Berru Mestanza50% (2)
- 6 Fisiología de La VisiónDocument49 pagini6 Fisiología de La VisiónManuelÎncă nu există evaluări
- Capitulo 18 FisiologiaDocument40 paginiCapitulo 18 FisiologiaBrenda Mota100% (1)
- Neurofisiologia Motora e IntegradoraDocument2 paginiNeurofisiologia Motora e IntegradoraSara Michell Astacio Rosales100% (1)
- Anatomia e Histología CardiovascularDocument6 paginiAnatomia e Histología CardiovascularTëfÿ DököÎncă nu există evaluări
- Informe S4-FISIOLOGÍA SEM-ES023 VVDocument15 paginiInforme S4-FISIOLOGÍA SEM-ES023 VVCoraine Chávez CórdovaÎncă nu există evaluări
- LCRDocument50 paginiLCRalejandry100% (4)
- Tema 2 Guia 2.2.Document3 paginiTema 2 Guia 2.2.Daniel Luis Meléndez ChoqueÎncă nu există evaluări
- Anatomía y Embriología de Órganos y SistemasDocument167 paginiAnatomía y Embriología de Órganos y SistemasCristóbal Muñoz Pérez100% (2)
- 03 Segmentacion MamiferosDocument8 pagini03 Segmentacion MamiferosMamen Casado MuñozÎncă nu există evaluări
- Desarrollo embrionario tempranoDocument20 paginiDesarrollo embrionario tempranoAlex Port0% (1)
- Desarrollo EmbrionarioDocument11 paginiDesarrollo EmbrionarioalejandravivianadaviÎncă nu există evaluări
- Tipos de HuevoDocument14 paginiTipos de HuevoAntonio Cortazar DiazÎncă nu există evaluări
- Desarrollo y reproducción en metazoosDocument13 paginiDesarrollo y reproducción en metazoosjuanma Giraldez MachucaÎncă nu există evaluări
- Desarrollo EmbrionarioDocument3 paginiDesarrollo EmbrionarioDiana SherlynÎncă nu există evaluări
- Etapasdeldesarrolloembrionario 120506113611 Phpapp01Document14 paginiEtapasdeldesarrolloembrionario 120506113611 Phpapp01Ricardo TapiaÎncă nu există evaluări
- Desarrollo Embrionario - Nataly Sofia Poma PerezDocument17 paginiDesarrollo Embrionario - Nataly Sofia Poma PerezAngie AguilarÎncă nu există evaluări
- Desarrollo Embrionario - Nataly Sofia Poma PerezDocument17 paginiDesarrollo Embrionario - Nataly Sofia Poma PerezAngie AguilarÎncă nu există evaluări
- Embriologia y AnatomiaDocument3 paginiEmbriologia y AnatomiaANABEL MAYÎncă nu există evaluări
- Reproducción biológica: Aspectos teóricosDocument26 paginiReproducción biológica: Aspectos teóricosMateoÎncă nu există evaluări
- Capitulo 15 BIOCELDocument44 paginiCapitulo 15 BIOCELLeonardo GuzmanÎncă nu există evaluări
- 03b SMITH EL CEREBRO PDFDocument89 pagini03b SMITH EL CEREBRO PDFmrchemistÎncă nu există evaluări
- Histología Cátedra 2 - TP 2Document1 paginăHistología Cátedra 2 - TP 2alexisdieserÎncă nu există evaluări
- Oxidación de Los Ácidos GrasosDocument37 paginiOxidación de Los Ácidos GrasosAída de AlonsoÎncă nu există evaluări
- Apunts Pem 1R Parcial PDFDocument452 paginiApunts Pem 1R Parcial PDFmanolo aceitunasÎncă nu există evaluări
- Alteración Genética en OrganismosDocument16 paginiAlteración Genética en OrganismosRivera Malaquías JuanÎncă nu există evaluări
- Folleto Trans Fobia 2015Document2 paginiFolleto Trans Fobia 2015KevinWilliams001Încă nu există evaluări
- Resistencia AntimicrobianaDocument3 paginiResistencia AntimicrobianaElvia RiveraÎncă nu există evaluări
- Banqueo BiologiaDocument94 paginiBanqueo Biologiabriyithe del pilar flores julcaÎncă nu există evaluări
- PRIMER PARCIAL - 23 - 09 - Revisión Del IntentoDocument11 paginiPRIMER PARCIAL - 23 - 09 - Revisión Del IntentoTamaraÎncă nu există evaluări
- Fases de La Mitosis Eva MeloDocument2 paginiFases de La Mitosis Eva MeloEva Flor Melo HernandezÎncă nu există evaluări
- Estrés en Adolescentes PDFDocument110 paginiEstrés en Adolescentes PDFCésar Hernández Lara100% (3)
- Talleres de Maturana Anexo19Document25 paginiTalleres de Maturana Anexo19maryruthcfÎncă nu există evaluări
- Nitazoxanida en El Tratamiento de La Diarrea Severa Por RotavirusDocument10 paginiNitazoxanida en El Tratamiento de La Diarrea Severa Por RotavirusshejurÎncă nu există evaluări
- Cuadro ComparativoDocument1 paginăCuadro ComparativoBraulio SánchezÎncă nu există evaluări
- Enzimas de RestriccionDocument10 paginiEnzimas de RestriccionYesika Paola Jaramillo14Încă nu există evaluări
- Clases 5 - 6 Biologia - CUI UNIVERSIDAD NACIONAL DE TUCUMANDocument7 paginiClases 5 - 6 Biologia - CUI UNIVERSIDAD NACIONAL DE TUCUMANClaudia CampanÎncă nu există evaluări
- Biologia Celular (BL) (BQ)Document3 paginiBiologia Celular (BL) (BQ)nigax XcÎncă nu există evaluări
- 1era Clase de Biología CelularDocument76 pagini1era Clase de Biología CelularMaria Belen MartinezÎncă nu există evaluări
- Neuromielitis OpticaDocument11 paginiNeuromielitis OpticaJorge De Vera AlvaradoÎncă nu există evaluări
- Las Plantas y Sus Partes DctoDocument4 paginiLas Plantas y Sus Partes DctoSilvana CabreraÎncă nu există evaluări
- Práctica 2 - Guia Practicas MoluscosDocument5 paginiPráctica 2 - Guia Practicas MoluscosIratxe SebastiánÎncă nu există evaluări
- Biolo 2Document21 paginiBiolo 2Rod Torres100% (1)
- Oiv Viti 564b 2019 EsDocument19 paginiOiv Viti 564b 2019 EsJesúsÎncă nu există evaluări
- Cuestionario Origen de La Vida Sobre La TierraDocument3 paginiCuestionario Origen de La Vida Sobre La TierraRigo Ochoa ArellanoÎncă nu există evaluări
- V. Ev 1 CatálogoDocument28 paginiV. Ev 1 Catálogojoselyne luqueÎncă nu există evaluări
- Informe Bioquimica IIDocument21 paginiInforme Bioquimica IIJuan Tarqui VelázquezÎncă nu există evaluări
- Informe de LaboratorioDocument63 paginiInforme de LaboratorioAura GuzmanÎncă nu există evaluări