S-ar putea să vă placă și
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Growing Canna Lilies in Your GardenDocument18 paginiGrowing Canna Lilies in Your GardenDennis CareyÎncă nu există evaluări
- 9 To 1 Balun For End Fed AntennaDocument24 pagini9 To 1 Balun For End Fed AntennaKlaudio KostaÎncă nu există evaluări
- Effect of Lanzones (Larum Domesticum) and Banana (Musa) Peel Fertilizer On Siling Labuyo (Capsicum Frutescens) PlantDocument3 paginiEffect of Lanzones (Larum Domesticum) and Banana (Musa) Peel Fertilizer On Siling Labuyo (Capsicum Frutescens) Plantsue lara espormaÎncă nu există evaluări
- Typology of Nursing Problems in Family Nursing PracticeDocument17 paginiTypology of Nursing Problems in Family Nursing PracticeAnonymous iG0DCOfÎncă nu există evaluări
- MaldivesDocument2 paginiMaldivesjemaripenyekÎncă nu există evaluări
- Secret Life of The Unborn ChildDocument3 paginiSecret Life of The Unborn ChildGreyzielle Gisella Buenafe TabulaÎncă nu există evaluări
- Secret Life of The Unborn ChildDocument3 paginiSecret Life of The Unborn ChildGreyzielle Gisella Buenafe TabulaÎncă nu există evaluări
- Secret Life of The Unborn ChildDocument3 paginiSecret Life of The Unborn ChildGreyzielle Gisella Buenafe TabulaÎncă nu există evaluări
- Rezolvari Subiecte Engleza Oral - BacDocument6 paginiRezolvari Subiecte Engleza Oral - BacteacherhelenÎncă nu există evaluări
- COPDDocument55 paginiCOPDRetno Lies SetiyowatiÎncă nu există evaluări
- 6048 FarbalDocument5 pagini6048 FarbalAryanto DedyÎncă nu există evaluări
- Research Article Studies On Transdermal Delivery Enhancement of ZidovudineDocument10 paginiResearch Article Studies On Transdermal Delivery Enhancement of ZidovudineAryanto DedyÎncă nu există evaluări
- 150cavity Synchronization by Extended Loop in Neuraminidase of 2009 h1n1 Influenza Type A Virus A Simulation StudyDocument11 pagini150cavity Synchronization by Extended Loop in Neuraminidase of 2009 h1n1 Influenza Type A Virus A Simulation StudyAryanto DedyÎncă nu există evaluări
- AbstractsDocument1 paginăAbstractsAryanto DedyÎncă nu există evaluări
- Research Article Studies On Transdermal Delivery Enhancement of ZidovudineDocument10 paginiResearch Article Studies On Transdermal Delivery Enhancement of ZidovudineAryanto DedyÎncă nu există evaluări
- HerlinaDocument1 paginăHerlinaAryanto DedyÎncă nu există evaluări
- Jandry KtiDocument1 paginăJandry KtiAryanto DedyÎncă nu există evaluări
- 17 Ifrj 20 (06) 2013 Adeleke 280Document13 pagini17 Ifrj 20 (06) 2013 Adeleke 280Aryanto DedyÎncă nu există evaluări
- 292-2010 SarkaDocument11 pagini292-2010 SarkaAryanto DedyÎncă nu există evaluări
- Clam Tmifsud PDFDocument0 paginiClam Tmifsud PDFAryanto DedyÎncă nu există evaluări
- Cell Surface Hydrophobicity, and Production of Proteinase and Hemolysin in Candida AlbicansDocument0 paginiCell Surface Hydrophobicity, and Production of Proteinase and Hemolysin in Candida AlbicansAryanto DedyÎncă nu există evaluări
- DepresipostpartumDocument13 paginiDepresipostpartumAryanto DedyÎncă nu există evaluări
- Jurnal FarmasiDocument18 paginiJurnal FarmasiAryanto DedyÎncă nu există evaluări
- Science Word SearchDocument1 paginăScience Word SearchHashimi IsmailÎncă nu există evaluări
- Reception Roster 1Document3 paginiReception Roster 1jassichadhaÎncă nu există evaluări
- Finals On Purposive Communication 2Document4 paginiFinals On Purposive Communication 2Domilyn Zantua BallaÎncă nu există evaluări
- Welcome Speech - 08 - Parbud Siang B - I Putu Adi Ardana YasaDocument4 paginiWelcome Speech - 08 - Parbud Siang B - I Putu Adi Ardana YasaAdi ArdanaÎncă nu există evaluări
- Price List Juni 2022 Plus GramasiDocument4 paginiPrice List Juni 2022 Plus GramasiElva ElvaÎncă nu există evaluări
- Pizza Hut PakistanDocument20 paginiPizza Hut PakistanMadiha KamranÎncă nu există evaluări
- Fish Meal - FAO Torry Research StationDocument10 paginiFish Meal - FAO Torry Research StationChiqui DomingoÎncă nu există evaluări
- HeinzDocument4 paginiHeinzShubham Ranka100% (1)
- Lactose Intolerance: C2 - Group 3Document29 paginiLactose Intolerance: C2 - Group 3Mark PadulloÎncă nu există evaluări
- Milk CigarDocument5 paginiMilk CigarMicha Andrea SanchezÎncă nu există evaluări
- Nef Preint Filetest 2bDocument7 paginiNef Preint Filetest 2bJorik VartanovÎncă nu există evaluări
- FACE TO FACE PRE INTERMEDIATE - Unit 5 TestDocument3 paginiFACE TO FACE PRE INTERMEDIATE - Unit 5 TestSonia GarciaÎncă nu există evaluări
- SUST 353 EXAM 2 Reading QsDocument20 paginiSUST 353 EXAM 2 Reading QsDaniel Amodeo-ChavezÎncă nu există evaluări
- QuestionPaper-Dairy ProcessingDocument1 paginăQuestionPaper-Dairy Processingharshitha jagathiesh100% (2)
- Unit 10A It Changed My Life 1 Grammar A)Document4 paginiUnit 10A It Changed My Life 1 Grammar A)Carmen GloriaÎncă nu există evaluări
- Joint Hindu Family Business - ModernisationDocument3 paginiJoint Hindu Family Business - Modernisationsimonebandrawala9100% (1)
- Lesson Presentation Types and Functions of TeethDocument16 paginiLesson Presentation Types and Functions of TeethNathan NwangumaÎncă nu există evaluări
- 1 Year New NotesDocument75 pagini1 Year New NotesPoornima Adithi100% (2)
- Espela History of Food StylingDocument1 paginăEspela History of Food StylingYohenyoÎncă nu există evaluări
- Street VendorsDocument26 paginiStreet VendorsBharat Bhushan SinghÎncă nu există evaluări
- Narrative Text Excercise SDocument12 paginiNarrative Text Excercise Srasjabrik18Încă nu există evaluări
- Tle Cookery Grade 10 Module 6Document17 paginiTle Cookery Grade 10 Module 6Lilac Qt100% (1)
- ch11Document27 paginich11tintfenÎncă nu există evaluări
- Indian Agrochemicals Industry: Insights and Outlook: February, 2019 I Industry ResearchDocument17 paginiIndian Agrochemicals Industry: Insights and Outlook: February, 2019 I Industry ResearchVenkatram PailaÎncă nu există evaluări
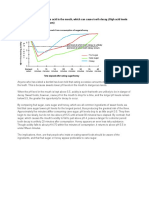
- Eating Sweet Foods Produces Acid in The Mouth, Which Can Cause Tooth Decay. (High Acid Levels Are Measured by Low PH Values)Document19 paginiEating Sweet Foods Produces Acid in The Mouth, Which Can Cause Tooth Decay. (High Acid Levels Are Measured by Low PH Values)Munawir Rahman IslamiÎncă nu există evaluări