S-ar putea să vă placă și
- Demystifying Viral Anterior Uveitis: A Review: Nicole Shu-Wen Chan Mbbs - Soon-Phaik Chee Frcophth Mmed (S'Pore)Document14 paginiDemystifying Viral Anterior Uveitis: A Review: Nicole Shu-Wen Chan Mbbs - Soon-Phaik Chee Frcophth Mmed (S'Pore)Dr Pranesh BalasubramaniamÎncă nu există evaluări
- Oftalmitis SimpatikaDocument12 paginiOftalmitis SimpatikaKarina HelsaÎncă nu există evaluări
- Frosted Branch AngiitisDocument9 paginiFrosted Branch AngiitisDragos-Constantin LuncaÎncă nu există evaluări
- Review: Ocular Toxoplasmosis II: Clinical Features, Pathology and ManagementDocument14 paginiReview: Ocular Toxoplasmosis II: Clinical Features, Pathology and Managementhidrisdmnk_747467972Încă nu există evaluări
- Dafpus 10 Sympathetic - Opthalmia-2Document7 paginiDafpus 10 Sympathetic - Opthalmia-2Anonymous jh87ryÎncă nu există evaluări
- The Basics of Uveitis: Goal StatementDocument13 paginiThe Basics of Uveitis: Goal StatementGreysia ManarisipÎncă nu există evaluări
- Immunopathology in Ocular Toxoplasmosis: Facts and Clues: Justus G Garweg /, Ermanno CandolfiDocument10 paginiImmunopathology in Ocular Toxoplasmosis: Facts and Clues: Justus G Garweg /, Ermanno CandolfiJuan Andrés Castaño CisfÎncă nu există evaluări
- Feline UveitesDocument9 paginiFeline UveitesErick ConfickeerzÎncă nu există evaluări
- Phacoanaphylactic Endophthalmitis: A Clinicopathologic ReviewDocument9 paginiPhacoanaphylactic Endophthalmitis: A Clinicopathologic ReviewLuci PonceÎncă nu există evaluări
- FinalDocument105 paginiFinalBharath BalagaÎncă nu există evaluări
- Systemic Disorders Associated With Peripheral Corneal UlcerationDocument4 paginiSystemic Disorders Associated With Peripheral Corneal UlcerationYoga KharismaÎncă nu există evaluări
- Hypopyon UveitisDocument18 paginiHypopyon UveitisjochigÎncă nu există evaluări
- Clinical Approach BlistersDocument25 paginiClinical Approach BlistersIsaacGonzalezÎncă nu există evaluări
- Retinal Vasculitis DDDocument17 paginiRetinal Vasculitis DDPopuri SandhyaÎncă nu există evaluări
- UveitisDocument45 paginiUveitistomeytto100% (1)
- Section 2: Diagnostic and Treatment Decisions: 2.32 Painful OphthalmoplegiaDocument14 paginiSection 2: Diagnostic and Treatment Decisions: 2.32 Painful Ophthalmoplegiamutya yulindaÎncă nu există evaluări
- Vasculitis ArticuloDocument22 paginiVasculitis Articuloelmer yohanÎncă nu există evaluări
- Seminars in Diagnostic Pathology: Mark R. WickDocument11 paginiSeminars in Diagnostic Pathology: Mark R. WickAbdillah AkbarÎncă nu există evaluări
- Uveitis GlaukomaDocument13 paginiUveitis GlaukomaDede FatmawatiÎncă nu există evaluări
- Uveitis 4Document9 paginiUveitis 4srihandayaniakbarÎncă nu există evaluări
- DR Vishali's PaperDocument9 paginiDR Vishali's PaperpoojasharmachdÎncă nu există evaluări
- Psoriasis and Uveitis A Literature ReviewDocument8 paginiPsoriasis and Uveitis A Literature Reviewc5kgkdd4100% (1)
- Periorbital Edema: A Puzzle No More?: ReviewDocument10 paginiPeriorbital Edema: A Puzzle No More?: ReviewJavier Infantes MolinaÎncă nu există evaluări
- AOP Krásný ENDocument9 paginiAOP Krásný ENDavid MartínezÎncă nu există evaluări
- Blau SyndDocument9 paginiBlau Synddianis0506Încă nu există evaluări
- KeratitisDocument26 paginiKeratitisHIDAYATUL RAHMIÎncă nu există evaluări
- Ocular Manifestations of SLEDocument9 paginiOcular Manifestations of SLEChiquitaPutriVaniaRauÎncă nu există evaluări
- Seminars in Diagnostic Pathology: Bullous, Pseudobullous, & Pustular DermatosesDocument11 paginiSeminars in Diagnostic Pathology: Bullous, Pseudobullous, & Pustular DermatosesCynthia OktariszaÎncă nu există evaluări
- Infectious Uveitis AyuDocument19 paginiInfectious Uveitis AyuAndi Ayu LestariÎncă nu există evaluări
- UveitisDocument11 paginiUveitisLuphly TaluvtaÎncă nu există evaluări
- Ocular Manifestations in Systemic Lupus Erythematosus: Sukhum Silpa-Archa, Joan J Lee, C Stephen FosterDocument8 paginiOcular Manifestations in Systemic Lupus Erythematosus: Sukhum Silpa-Archa, Joan J Lee, C Stephen FosterNadya Regina PermataÎncă nu există evaluări
- Okap Samson Uveitis PDFDocument12 paginiOkap Samson Uveitis PDFsharu4291Încă nu există evaluări
- Major Review: Glaucoma and UveitisDocument10 paginiMajor Review: Glaucoma and UveitisAlex MuñozÎncă nu există evaluări
- HMR 36 155 PDFDocument6 paginiHMR 36 155 PDFFatimatus Zahroh IchsanÎncă nu există evaluări
- Anterior ÜveitDocument11 paginiAnterior Üveitmuhammeteminkaya914Încă nu există evaluări
- Seasonal Monthly Variation Amongst Reported Cataract Surgeries in IndiaDocument92 paginiSeasonal Monthly Variation Amongst Reported Cataract Surgeries in IndiafriendsofindiaÎncă nu există evaluări
- 10.1007@s10792 019 01183 9Document8 pagini10.1007@s10792 019 01183 9adriana gutanuÎncă nu există evaluări
- Vasculitis LeucocitoclásticaDocument11 paginiVasculitis LeucocitoclásticaLaura Daniela MurilloÎncă nu există evaluări
- Welder Et Al (2012) - Herpes Simplex Keratitis. Available At: Accessed On: 20 Agustus 2015Document24 paginiWelder Et Al (2012) - Herpes Simplex Keratitis. Available At: Accessed On: 20 Agustus 2015Silvestri PurbaÎncă nu există evaluări
- Sympathetic Uveitis/ Ophthalmia: PG CornerDocument4 paginiSympathetic Uveitis/ Ophthalmia: PG CornerVincent LivandyÎncă nu există evaluări
- Episcleritis - UpToDateDocument15 paginiEpiscleritis - UpToDateanisa maria ulfaÎncă nu există evaluări
- Proceeding of The SEVC Southern European Veterinary ConferenceDocument4 paginiProceeding of The SEVC Southern European Veterinary ConferenceHartina SamosirÎncă nu există evaluări
- Articulo Uveitis Anterior ViralDocument10 paginiArticulo Uveitis Anterior ViralJohana DíazÎncă nu există evaluări
- Thyroid Eye Disease: Autoimmune Disorder Characterised by Infiltrative OrbitopathyDocument43 paginiThyroid Eye Disease: Autoimmune Disorder Characterised by Infiltrative OrbitopathyBIBI MOHANANÎncă nu există evaluări
- Equine Recurrent Uveitis: U.S.A.M.V. Cluj-Napoca Facultatea de Medicină Veterinară Disciplina: OftalmologieDocument9 paginiEquine Recurrent Uveitis: U.S.A.M.V. Cluj-Napoca Facultatea de Medicină Veterinară Disciplina: OftalmologiePatricia PopÎncă nu există evaluări
- AVIAN - Diseases of The Avian SpleenDocument7 paginiAVIAN - Diseases of The Avian Spleentaner_soysuren100% (1)
- Herpes Zoster Ophthalmicus Review and Prevention: Andrew R. Davis, and John SheppardDocument6 paginiHerpes Zoster Ophthalmicus Review and Prevention: Andrew R. Davis, and John SheppardVincentius Michael WilliantoÎncă nu există evaluări
- Glaucoma Associated With UveitisDocument34 paginiGlaucoma Associated With UveitisHy Vui VeÎncă nu există evaluări
- Ocular Allergy Esen Karamursel Akpek, MDDocument8 paginiOcular Allergy Esen Karamursel Akpek, MDAndri SaputraÎncă nu există evaluări
- Alergia em CaesDocument17 paginiAlergia em CaesNícolas AugustoÎncă nu există evaluări
- Prepared by Getenet Shumet (BSC, MSC Student)Document64 paginiPrepared by Getenet Shumet (BSC, MSC Student)Getenet shumetÎncă nu există evaluări
- Clinical Features and Treatment of Ocular Toxoplasmosis: AbstractDocument8 paginiClinical Features and Treatment of Ocular Toxoplasmosis: AbstractInne Budi Kusuma WardhaniÎncă nu există evaluări
- Presumed Ocular Histoplasmosis: ReviewDocument5 paginiPresumed Ocular Histoplasmosis: ReviewDiana PSÎncă nu există evaluări
- Viral Posterior UveitisDocument42 paginiViral Posterior UveitisJavier Infantes MolinaÎncă nu există evaluări
- Seminars in Diagnostic Pathology: Mark R. WickDocument12 paginiSeminars in Diagnostic Pathology: Mark R. WickNancy ManriqueÎncă nu există evaluări
- EPISCLERITIS & ScleritisDocument14 paginiEPISCLERITIS & ScleritisChikita Artia SariÎncă nu există evaluări
- Conjunctivitis, Allergic: Author: Parag A Majmudar, MD, Fellowship Co-Director, Department of OphthalmologyDocument7 paginiConjunctivitis, Allergic: Author: Parag A Majmudar, MD, Fellowship Co-Director, Department of OphthalmologyDicki Pratama HolmesÎncă nu există evaluări
- Eye Docs UveitisDocument90 paginiEye Docs UveitisMuneeb ShahzadÎncă nu există evaluări
- Mamen SisDocument15 paginiMamen SisIwa Fathi SyahdiaÎncă nu există evaluări
- Teamwork For Better Collaboration Additional Reading Material PDFDocument7 paginiTeamwork For Better Collaboration Additional Reading Material PDFMyScribd_ieltsÎncă nu există evaluări
- Mask Making and Make A CardDocument2 paginiMask Making and Make A CardMyScribd_ieltsÎncă nu există evaluări
- Mhnraopymntgldprcse 16092020 PDFDocument1 paginăMhnraopymntgldprcse 16092020 PDFMyScribd_ieltsÎncă nu există evaluări
- Fractions: More Questions For PracticeDocument2 paginiFractions: More Questions For PracticeMyScribd_ieltsÎncă nu există evaluări
- Physical Quantities and Measurement STD 7 Physics NotesDocument6 paginiPhysical Quantities and Measurement STD 7 Physics NotesMyScribd_ieltsÎncă nu există evaluări
- 12 Tips For Troubleshooting Your Internet ConnectionDocument12 pagini12 Tips For Troubleshooting Your Internet ConnectionMyScribd_ieltsÎncă nu există evaluări
- I Keep Getting A Message That Says "Your Machine Administrator Disabled Teredo Locally."Document4 paginiI Keep Getting A Message That Says "Your Machine Administrator Disabled Teredo Locally."MyScribd_ieltsÎncă nu există evaluări
- Bhagavad-Gita Dictionary PDFDocument128 paginiBhagavad-Gita Dictionary PDFMyScribd_ieltsÎncă nu există evaluări
- Tales of Arjuna PDFDocument34 paginiTales of Arjuna PDFMyScribd_ieltsÎncă nu există evaluări
- Clever Rabbit and Bear Goes FishingDocument2 paginiClever Rabbit and Bear Goes FishingMyScribd_ieltsÎncă nu există evaluări
- How To Learn Python For Data ScienceDocument22 paginiHow To Learn Python For Data ScienceMyScribd_ieltsÎncă nu există evaluări
- Netsh Command SyntaxDocument6 paginiNetsh Command SyntaxMyScribd_ieltsÎncă nu există evaluări
- Analysis and Presentation For Bank Marketing Data: Vinay Kumar MS by Research Scholar IIT Kharagpur +91-8348575432Document20 paginiAnalysis and Presentation For Bank Marketing Data: Vinay Kumar MS by Research Scholar IIT Kharagpur +91-8348575432MyScribd_ieltsÎncă nu există evaluări
- Anatella Tutorial BasicDocument41 paginiAnatella Tutorial BasicMyScribd_ielts100% (1)
- Math For Data ScienceDocument11 paginiMath For Data ScienceMyScribd_ieltsÎncă nu există evaluări
- Disease Review 1-53 New 5 MayDocument50 paginiDisease Review 1-53 New 5 MayMyScribd_ieltsÎncă nu există evaluări
- Teradata SyllabusDocument3 paginiTeradata SyllabusMyScribd_ieltsÎncă nu există evaluări
- Ophthalmology Easy GuideDocument46 paginiOphthalmology Easy Guidesri sinaga0% (1)
- Eye Diagnostic Points - McqsDocument21 paginiEye Diagnostic Points - McqsMuhammad AwaisÎncă nu există evaluări
- Jurnal UveitisDocument5 paginiJurnal UveitisAnna ApsariÎncă nu există evaluări
- Red Eye - Conjunctivitis I Dr. Nuke Erlina Mayasari, SPMDocument23 paginiRed Eye - Conjunctivitis I Dr. Nuke Erlina Mayasari, SPMismkipendprowil2Încă nu există evaluări
- AAO CatalogDocument80 paginiAAO CatalogLetchumana Krishnan50% (2)
- Standardization of Uveitis Nomenclature For Reporting Clinical D 2005Document8 paginiStandardization of Uveitis Nomenclature For Reporting Clinical D 2005Stephanie PfengÎncă nu există evaluări
- GlaucomaDocument53 paginiGlaucomaLyÎncă nu există evaluări
- Referat Vogt-Koyanagi-Harada Disease Vitha (Revisi)Document41 paginiReferat Vogt-Koyanagi-Harada Disease Vitha (Revisi)vitha pranamaÎncă nu există evaluări
- Signs and Symptom For UveitisDocument4 paginiSigns and Symptom For UveitisVan KochkarianÎncă nu există evaluări
- Casos Clínicos Sobre UveíteDocument16 paginiCasos Clínicos Sobre UveíteMirella Lopes da RochaÎncă nu există evaluări
- PRIMADocument192 paginiPRIMAKilluaZoldyckÎncă nu există evaluări
- Eye ImmunologyDocument35 paginiEye ImmunologySingo DemurÎncă nu există evaluări
- Non-Infectious (NI) Intermediate, Posterior and Panuveitis ICD CodesDocument2 paginiNon-Infectious (NI) Intermediate, Posterior and Panuveitis ICD CodesSiswoÎncă nu există evaluări
- 2017 Tubulointerstitial Nephritis, Diagnosis, Treatment, and MonitoringDocument11 pagini2017 Tubulointerstitial Nephritis, Diagnosis, Treatment, and MonitoringbrufenÎncă nu există evaluări
- MataDocument16 paginiMataIslah NadilaÎncă nu există evaluări
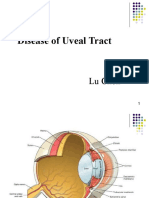
- Diseases of Uveal TractDocument62 paginiDiseases of Uveal TractIhsan HanifÎncă nu există evaluări
- 100 Opthalmology Questions For PracticeDocument37 pagini100 Opthalmology Questions For Practicekyonaboy85% (13)
- Cystoid Macular EdemaDocument5 paginiCystoid Macular EdemaAgnes Triana BasjaÎncă nu există evaluări
- Notes in Ophthalmology: MCQ, Osce, SlidDocument21 paginiNotes in Ophthalmology: MCQ, Osce, SlidDrmhdh DrmhdhÎncă nu există evaluări
- Uveit FosterDocument954 paginiUveit FosterEdmond IsufajÎncă nu există evaluări
- Eye Mcqs 1.Document11 paginiEye Mcqs 1.muhammad kazimÎncă nu există evaluări
- Glaucoma, Hyphema: Synonyms and Related Keywords: Hyphema, Microhyphema, Hemorrhage in The AnteriorDocument22 paginiGlaucoma, Hyphema: Synonyms and Related Keywords: Hyphema, Microhyphema, Hemorrhage in The AnteriorWulandari EmyÎncă nu există evaluări
- Uveitis WilliamDocument159 paginiUveitis WilliamSaul Ricardo Zambrano OyagueÎncă nu există evaluări
- LE5 ChecklistDocument2 paginiLE5 ChecklistBrendi MarzanÎncă nu există evaluări
- Posterior ScleritisDocument4 paginiPosterior ScleritisPande GustianaÎncă nu există evaluări
- AAO UveitisDocument39 paginiAAO UveitisSafa Abdualrahaman Ali HamadÎncă nu există evaluări
- Ocular Hypotony - A Comprehensive ReviewDocument20 paginiOcular Hypotony - A Comprehensive ReviewJavier Infantes MolinaÎncă nu există evaluări
- Ophthalmoology Study Notes ComprehensiveDocument97 paginiOphthalmoology Study Notes ComprehensivekabeerismailÎncă nu există evaluări
- 9 Samuel EtalDocument4 pagini9 Samuel EtaleditorijmrhsÎncă nu există evaluări
- AIIMS PG Solved Paper 2001Document23 paginiAIIMS PG Solved Paper 2001shaffeque0% (1)