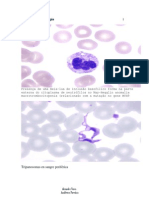
S-ar putea să vă placă și
- Ante Project oDocument135 paginiAnte Project onsadisi_195165533Încă nu există evaluări
- CatalogoGeral Ensitel 2013Document38 paginiCatalogoGeral Ensitel 2013Valódia VictorinoÎncă nu există evaluări
- Aquario Agua Doce - Curso CompletoDocument25 paginiAquario Agua Doce - Curso Completocucick100% (1)
- Como Manter Saudáveis Os Seus Peixes OrnamentaisDocument55 paginiComo Manter Saudáveis Os Seus Peixes Ornamentaisadatti100% (1)
- Como Economizarnas Comprasem SupermercadosDocument0 paginiComo Economizarnas Comprasem SupermercadosValódia VictorinoÎncă nu există evaluări
- Aquario Agua Doce - Curso CompletoDocument25 paginiAquario Agua Doce - Curso Completocucick100% (1)
- Plano Ensino Comportamento Organizacional PDFDocument14 paginiPlano Ensino Comportamento Organizacional PDFValódia VictorinoÎncă nu există evaluări
- Monte Seu Plano Nut Ric IonalDocument0 paginiMonte Seu Plano Nut Ric IonalValódia VictorinoÎncă nu există evaluări
- Abdomen Trin CA DoDocument17 paginiAbdomen Trin CA DofabriziodragonÎncă nu există evaluări
- Guia de Nutri CaoDocument0 paginiGuia de Nutri CaoValódia VictorinoÎncă nu există evaluări
- Kinguio: pacífico e colorido peixe ornamentalDocument7 paginiKinguio: pacífico e colorido peixe ornamentalValódia VictorinoÎncă nu există evaluări
- Caracteristicas Da Central de IncendioDocument2 paginiCaracteristicas Da Central de IncendioValódia VictorinoÎncă nu există evaluări
- Coca ColaDocument1 paginăCoca ColaValódia VictorinoÎncă nu există evaluări
- Livro Bimby MassasDocument3 paginiLivro Bimby MassasValódia VictorinoÎncă nu există evaluări
- Coca ColaDocument1 paginăCoca ColaValódia VictorinoÎncă nu există evaluări
- Prova de biologia sobre células, fotossíntese e respiraçãoDocument3 paginiProva de biologia sobre células, fotossíntese e respiraçãoNathalia LopesÎncă nu există evaluări
- AULA - ALIMENTAÇAO ANIMAL - SLIDEpptxDocument24 paginiAULA - ALIMENTAÇAO ANIMAL - SLIDEpptxLourenço ChiteteÎncă nu există evaluări
- Atlas Hematologia Citipatologia 1Document40 paginiAtlas Hematologia Citipatologia 1Luciano Francisco100% (2)
- Reprodução das cecílias: modos reprodutivos e cuidados parentaisDocument2 paginiReprodução das cecílias: modos reprodutivos e cuidados parentaisIonara NunesÎncă nu există evaluări
- Roteiro Mensal - 3º Ano - FEVEREIRO-MARÇODocument2 paginiRoteiro Mensal - 3º Ano - FEVEREIRO-MARÇOGiuliane SantosÎncă nu există evaluări
- Magali Milene - A Saúde Mental e A Fabricação Da NormalidadeDocument10 paginiMagali Milene - A Saúde Mental e A Fabricação Da NormalidadeHarrison RivelloÎncă nu există evaluări
- Sociedade Primitiva PDFDocument20 paginiSociedade Primitiva PDFArquimedes AtlasÎncă nu există evaluări
- Sistema Nervoso MiotentericoDocument20 paginiSistema Nervoso MiotentericoJorge CamargoÎncă nu există evaluări
- Ecologia humana e sociabilidade urbanaDocument36 paginiEcologia humana e sociabilidade urbanadueduardaÎncă nu există evaluări
- Trocas Nutricionais Entre o Organismo Dos Animais e o MeioDocument5 paginiTrocas Nutricionais Entre o Organismo Dos Animais e o MeioPaula MonteiroÎncă nu există evaluări
- Sistema digestório: órgãos e funções emDocument29 paginiSistema digestório: órgãos e funções emRose Meire Oliveira BarrosÎncă nu există evaluări
- CIANOBACTÉRIASDocument3 paginiCIANOBACTÉRIASprofisioÎncă nu există evaluări
- Site BiofísicaDocument8 paginiSite BiofísicaLuciclaudio Silva BarbosaÎncă nu există evaluări
- Atividades de Fixação Da Primeira Lei de MendelDocument4 paginiAtividades de Fixação Da Primeira Lei de MendelChiara Cristina Marafon Correia100% (1)
- Histologia - HemocipoieseDocument62 paginiHistologia - HemocipoieseMarcelle MarquesÎncă nu există evaluări
- Microagulhamento Como Recurso de Tratamento NoDocument10 paginiMicroagulhamento Como Recurso de Tratamento NoIasmine PedrosoÎncă nu există evaluări
- Farmacocinética e formas de administração de medicamentosDocument59 paginiFarmacocinética e formas de administração de medicamentosnatalia100% (2)
- Ce 422327 PDFDocument4 paginiCe 422327 PDFGeruza De BritoÎncă nu există evaluări
- Drosophila MelanogasterDocument27 paginiDrosophila Melanogastermargaridasoares12Încă nu există evaluări
- Endogamia vs Heterose: conceitos e implicações em melhoramento de plantasDocument24 paginiEndogamia vs Heterose: conceitos e implicações em melhoramento de plantasEuclidesdaSilvaÎncă nu există evaluări
- O que é Educação FísicaDocument23 paginiO que é Educação FísicaJuan Bueno100% (1)
- Celulite: causas, sintomas e tratamentosDocument29 paginiCelulite: causas, sintomas e tratamentosJemima Daniela Dias MoraesÎncă nu există evaluări
- Sistema Imunitário SecundárioDocument5 paginiSistema Imunitário SecundárioMaria Maia100% (1)
- EXercícios Sobre EcologiaDocument8 paginiEXercícios Sobre EcologiaMágdaSales100% (1)
- Virus e Viroses - 3º Ano - Bio - 01Document58 paginiVirus e Viroses - 3º Ano - Bio - 01Igor CajatyÎncă nu există evaluări
- Biologia CAA 2s Vol2 2010reduzidoDocument48 paginiBiologia CAA 2s Vol2 2010reduzidoRones Dias100% (1)
- Meiose: processo de formação de gametas com metade do número cromossômicoDocument9 paginiMeiose: processo de formação de gametas com metade do número cromossômicoGabii FachinelloÎncă nu există evaluări
- Alguns Órgãos Que Compõem o Sistema Reprodutor Masculino SãoDocument9 paginiAlguns Órgãos Que Compõem o Sistema Reprodutor Masculino SãoViviane PiresÎncă nu există evaluări
- Plano de Aula AlgasDocument7 paginiPlano de Aula AlgasDanielle Diniz GalvãoÎncă nu există evaluări
- Exercícios - CiênciasDocument2 paginiExercícios - CiênciasMonica AlvesÎncă nu există evaluări