S-ar putea să vă placă și
- Paediatric AsthmaDocument246 paginiPaediatric AsthmaOxana TurcuÎncă nu există evaluări
- Paediatrics and Adult Immune SystemDocument14 paginiPaediatrics and Adult Immune SystemOxana TurcuÎncă nu există evaluări
- Pain Management in Infants - Children - AdolescentsDocument9 paginiPain Management in Infants - Children - AdolescentsOxana Turcu100% (1)
- Allergic Bronchopulmonary Aspergillosis Clinical Care Guidelines - CF FoundationDocument8 paginiAllergic Bronchopulmonary Aspergillosis Clinical Care Guidelines - CF FoundationOxana TurcuÎncă nu există evaluări
- Acute Exacerbations of Pulmonary DiseasesDocument258 paginiAcute Exacerbations of Pulmonary DiseasesOxana TurcuÎncă nu există evaluări
- Dalhousie Dyspnea ScalesDocument7 paginiDalhousie Dyspnea ScalesOxana TurcuÎncă nu există evaluări
- Anitbiotic Treatment in Cystic FibrosisDocument76 paginiAnitbiotic Treatment in Cystic FibrosisOxana TurcuÎncă nu există evaluări
- Down Syndrome Cognitive DisabilityDocument2 paginiDown Syndrome Cognitive DisabilityOxana TurcuÎncă nu există evaluări
- InTech-Arrhythmias in Children and Young AdultsDocument36 paginiInTech-Arrhythmias in Children and Young Adultsmadalina8Încă nu există evaluări
- Radiological Features of BronchiectasisDocument24 paginiRadiological Features of BronchiectasisOxana TurcuÎncă nu există evaluări
- Pediatric Emergencies PDFDocument20 paginiPediatric Emergencies PDFOxana Turcu100% (1)
- Peptic Ulcer Disease in Children PDFDocument9 paginiPeptic Ulcer Disease in Children PDFOxana TurcuÎncă nu există evaluări
- Clinical ExaminationDocument25 paginiClinical ExaminationOxana TurcuÎncă nu există evaluări
- The Pediatric History and Physical Exam PDFDocument18 paginiThe Pediatric History and Physical Exam PDFOxana TurcuÎncă nu există evaluări
- Biliary Dyskinesia PDFDocument3 paginiBiliary Dyskinesia PDFOxana TurcuÎncă nu există evaluări
- Bousquet PDFDocument11 paginiBousquet PDFOxana TurcuÎncă nu există evaluări
- Respiratory Failure in Children PDFDocument47 paginiRespiratory Failure in Children PDFOxana TurcuÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Retts SyndromeDocument10 paginiRetts Syndromeapi-297523122Încă nu există evaluări
- Neuroepigenomics in Aging and Disease PDFDocument520 paginiNeuroepigenomics in Aging and Disease PDFManoel CordeiroÎncă nu există evaluări
- Evaluation of The Child With Global Developmental DelayDocument14 paginiEvaluation of The Child With Global Developmental DelaylauramwoodyardÎncă nu există evaluări
- Athens 2010 IALP - PROCEEDINGS PDFDocument882 paginiAthens 2010 IALP - PROCEEDINGS PDFYelly Andriani Barlian100% (1)
- Prenatal ScreningDocument44 paginiPrenatal ScreningHoopmen tampubolonÎncă nu există evaluări
- Rett SyndromeDocument13 paginiRett SyndromeAileish Kate Lanuevo JaudianÎncă nu există evaluări
- Rett Syndrome 1Document19 paginiRett Syndrome 1api-545157726Încă nu există evaluări
- Autism A Neuroepigenetic DisorderDocument13 paginiAutism A Neuroepigenetic DisorderMelania TonelloÎncă nu există evaluări
- Thesis TitlesDocument25 paginiThesis TitlesMicah Dianne DizonÎncă nu există evaluări
- Chromatin RemodellingDocument234 paginiChromatin Remodellingplastioid4079Încă nu există evaluări
- Dna Methylation in Human Diseases: SciencedirectDocument8 paginiDna Methylation in Human Diseases: SciencedirectKarim KACEMÎncă nu există evaluări
- What Is Rett Syndrome?Document4 paginiWhat Is Rett Syndrome?Jon_NamikazeÎncă nu există evaluări
- Epigenetic Mechanisms in Developmental Alcohol Induceds Neurobehavioral Deficits DivididoDocument17 paginiEpigenetic Mechanisms in Developmental Alcohol Induceds Neurobehavioral Deficits DivididoJOHANNA CATHERINE RUIZ CASTILLOÎncă nu există evaluări
- Intellectual Disability in Children - Evaluation For A Cause - UpToDateDocument30 paginiIntellectual Disability in Children - Evaluation For A Cause - UpToDatecapt_zoe100% (1)
- Antipurinergic Therapy For Autism-An In-Depth Review.Document15 paginiAntipurinergic Therapy For Autism-An In-Depth Review.Miguel Romero100% (1)
- FDAapproves Daybuethefirsttreatmentfor RettsyndromeDocument8 paginiFDAapproves Daybuethefirsttreatmentfor RettsyndromeNeethu Anna StephenÎncă nu există evaluări
- Chromatin RemodellingDocument234 paginiChromatin RemodellingBetsabé Vega AbadÎncă nu există evaluări
- Abstracts From ESMED Congress 2021Document197 paginiAbstracts From ESMED Congress 2021European Society of Medicine (ESMED)100% (2)
- Science:, 792 (2003) Robin D. Rogers and Kenneth R. SeddonDocument3 paginiScience:, 792 (2003) Robin D. Rogers and Kenneth R. SeddonJohnSmithÎncă nu există evaluări
- The Role of Neuroglia in AutismDocument30 paginiThe Role of Neuroglia in AutismWesley M.SantosÎncă nu există evaluări
- Pediatric Neurology - Lawson Peter N. (SRG)Document200 paginiPediatric Neurology - Lawson Peter N. (SRG)Hriday DeÎncă nu există evaluări
- Autism Spectrum DisordersDocument9 paginiAutism Spectrum DisordersClaudia Verónica Revilla PazÎncă nu există evaluări
- GABAA Receptor Downregulation in Brains of Subjects With AutismDocument7 paginiGABAA Receptor Downregulation in Brains of Subjects With Autismaini qurrotullainÎncă nu există evaluări
- (Understanding Health and Sickness Series) M.D. Patricia Ainsworth, Ph.D. Pamela C. Baker-Understanding Mental Retardation (Understanding Health and Sickness Series) - University Press of MississippiDocument212 pagini(Understanding Health and Sickness Series) M.D. Patricia Ainsworth, Ph.D. Pamela C. Baker-Understanding Mental Retardation (Understanding Health and Sickness Series) - University Press of MississippiFajar YuniftiadiÎncă nu există evaluări
- Evaluation of The Child With Global Developmental DelayDocument14 paginiEvaluation of The Child With Global Developmental DelayplaincircleÎncă nu există evaluări
- Combinepdf 7Document129 paginiCombinepdf 7Joe JosephÎncă nu există evaluări
- Genetic Causes of Syndromic and Non-Syndromic Autism PDFDocument9 paginiGenetic Causes of Syndromic and Non-Syndromic Autism PDFRaisa CoppolaÎncă nu există evaluări
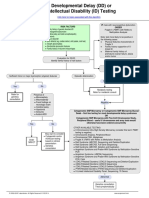
- Developmental Delay (DD) or Intellectual Disability (ID) Testing AlgorithmDocument1 paginăDevelopmental Delay (DD) or Intellectual Disability (ID) Testing AlgorithmNeo Mervyn Monaheng100% (1)
- The Role of Epigenetics in Mental Disorders: Review ArticleDocument7 paginiThe Role of Epigenetics in Mental Disorders: Review ArticleBenet DhasÎncă nu există evaluări
- Childhood DisordersDocument21 paginiChildhood DisordersstephanieÎncă nu există evaluări