S-ar putea să vă placă și
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Young FrankensteinDocument109 paginiYoung FrankensteinColleen Hawks-Pierce83% (6)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- 6th Central Pay Commission Salary CalculatorDocument15 pagini6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Obsessive-Compulsive Disorder PDFDocument6 paginiObsessive-Compulsive Disorder PDFSetsuna F Seiei100% (1)
- Psychological Assessment ReviewerDocument9 paginiPsychological Assessment Reviewerpatquianzon100% (1)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- Abdominal Aortic AneurysmDocument20 paginiAbdominal Aortic AneurysmPortia Rose RodriguezÎncă nu există evaluări
- Framingham Risk Score SaDocument8 paginiFramingham Risk Score Saapi-301624030Încă nu există evaluări
- Emotional Causes of CancerDocument6 paginiEmotional Causes of CancerstudiocrocÎncă nu există evaluări
- Miyawaki Yuki Kuwahara Koei Hari Handbook of Traditional JapDocument23 paginiMiyawaki Yuki Kuwahara Koei Hari Handbook of Traditional Japкостя100% (1)
- Rife RatesDocument17 paginiRife RateszonetrekÎncă nu există evaluări
- 4 Measurement and ComputationDocument6 pagini4 Measurement and ComputationBernardMarkMateoÎncă nu există evaluări
- Landes .Off - Pump.coronary - Artery.bypass - Surgery. 2005Document163 paginiLandes .Off - Pump.coronary - Artery.bypass - Surgery. 2005Setsuna F SeieiÎncă nu există evaluări
- Training of Top US Distance SwimmersDocument45 paginiTraining of Top US Distance SwimmersMel HarbourÎncă nu există evaluări
- GAME COLLECTIONDocument29 paginiGAME COLLECTIONSetsuna F SeieiÎncă nu există evaluări
- Differences between playthroughs in Sengoku RanceDocument101 paginiDifferences between playthroughs in Sengoku RanceSetsuna F SeieiÎncă nu există evaluări
- Jurnal Pneumothorax PDFDocument5 paginiJurnal Pneumothorax PDFSetsuna F SeieiÎncă nu există evaluări
- Obsessive-Compulsive Disorder Workup PDFDocument3 paginiObsessive-Compulsive Disorder Workup PDFSetsuna F SeieiÎncă nu există evaluări
- Principles of Diagnosis and Antimicrobial TherapyDocument116 paginiPrinciples of Diagnosis and Antimicrobial TherapyD'nollas LadysÎncă nu există evaluări
- Vital Signs at Various AgesDocument2 paginiVital Signs at Various AgesSetsuna F SeieiÎncă nu există evaluări
- Obsessive-Compulsive Disorder Clinical PresentationDocument4 paginiObsessive-Compulsive Disorder Clinical PresentationSetsuna F SeieiÎncă nu există evaluări
- Obsessive-Compulsive Disorder Treatment & Management PDFDocument4 paginiObsessive-Compulsive Disorder Treatment & Management PDFSetsuna F SeieiÎncă nu există evaluări
- GAME COLLECTIONDocument29 paginiGAME COLLECTIONSetsuna F SeieiÎncă nu există evaluări
- FF1 MatoyaDocument4 paginiFF1 MatoyaAlejandro TrinidadÎncă nu există evaluări
- Anatomi Medulla SpinalisDocument4 paginiAnatomi Medulla SpinalisOdineArgusÎncă nu există evaluări
- Current Clinical Strategies, Handbook of Psychiatric Drugs (2005) - BM OCR 7.0-2.5Document68 paginiCurrent Clinical Strategies, Handbook of Psychiatric Drugs (2005) - BM OCR 7.0-2.5Adina VarzaruÎncă nu există evaluări
- 9 Autophagy-Related Proteins, LC3 and Beclin-1, in Placentas From Pregnancies Complicated by PreeclmapsiaDocument10 pagini9 Autophagy-Related Proteins, LC3 and Beclin-1, in Placentas From Pregnancies Complicated by PreeclmapsiaSetsuna F SeieiÎncă nu există evaluări
- Bakul-Patofisiologi Trauma (Compatibility Mode)Document9 paginiBakul-Patofisiologi Trauma (Compatibility Mode)Setsuna F SeieiÎncă nu există evaluări
- Estimasi Kehilangan DarahDocument2 paginiEstimasi Kehilangan DarahSetsuna F SeieiÎncă nu există evaluări
- ZTE Service CenterDocument2 paginiZTE Service CenterSetsuna F SeieiÎncă nu există evaluări
- 03.anemia of Chronic DiseaseDocument6 pagini03.anemia of Chronic DiseaseYockie Verinico SimamoraÎncă nu există evaluări
- Prof. Soetamto-Blood Loss Estimation and Transfusion inDocument11 paginiProf. Soetamto-Blood Loss Estimation and Transfusion inElsya ParamitasariÎncă nu există evaluări
- Aplastic Anemia: Causes, Symptoms and TreatmentDocument16 paginiAplastic Anemia: Causes, Symptoms and TreatmentSetsuna F SeieiÎncă nu există evaluări
- SURAT KETERANGAN DOKTERDocument1 paginăSURAT KETERANGAN DOKTERalha egaÎncă nu există evaluări
- Continuous Passive Motion (CPM) : Theory and Principles of Clinical ApplicationDocument10 paginiContinuous Passive Motion (CPM) : Theory and Principles of Clinical ApplicationlaurentiaÎncă nu există evaluări
- Mink Dissection InstructionsDocument15 paginiMink Dissection InstructionsMark PenticuffÎncă nu există evaluări
- Aos 94 10Document11 paginiAos 94 10Andi Tiara S. AdamÎncă nu există evaluări
- ASPEN Pocket Cards - Guidelines For The Provision and Assessment of Nutrition Support Therapy in The Pediatric Critically Ill Patient-1Document12 paginiASPEN Pocket Cards - Guidelines For The Provision and Assessment of Nutrition Support Therapy in The Pediatric Critically Ill Patient-1Ceren KamalıÎncă nu există evaluări
- Latest Guideline on Caries-risk Assessment and ManagementDocument8 paginiLatest Guideline on Caries-risk Assessment and ManagementPriscila Belén Chuhuaicura SotoÎncă nu există evaluări
- Huntingtons Disease ProjectDocument9 paginiHuntingtons Disease Projectapi-306544419Încă nu există evaluări
- Notice Application Form For SRDocument4 paginiNotice Application Form For SRPankaj PrasadÎncă nu există evaluări
- 10 Furunculosis (Aeromonas Salmonicida)Document86 pagini10 Furunculosis (Aeromonas Salmonicida)Jose AntonioÎncă nu există evaluări
- Phagocytosis Assay Kit (Igg Fitc) : Item No. 500290Document9 paginiPhagocytosis Assay Kit (Igg Fitc) : Item No. 500290drfiatÎncă nu există evaluări
- Overdentures Improve Chewing Efficiency and Bone SupportDocument8 paginiOverdentures Improve Chewing Efficiency and Bone SupportMezo SalahÎncă nu există evaluări
- Q4 Health 6 Week3Document4 paginiQ4 Health 6 Week3Ethelyn Joy FarquerabaoÎncă nu există evaluări
- Aims and Applications: Aims: The Three Aims of Pharmacoepidemiology AreDocument10 paginiAims and Applications: Aims: The Three Aims of Pharmacoepidemiology Aregowtham0% (1)
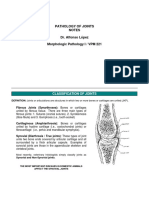
- Pathology of JointsDocument13 paginiPathology of JointswobblegobbleÎncă nu există evaluări
- Goblet CellDocument4 paginiGoblet CellMuhammad ZairulfikriÎncă nu există evaluări
- Assessment Diagnosis Planning Interventions Rationale Evaluation Subjective: "Maglisod Man Kog Short Term: Independent: - Establish RapportDocument3 paginiAssessment Diagnosis Planning Interventions Rationale Evaluation Subjective: "Maglisod Man Kog Short Term: Independent: - Establish RapportSergi Lee OrateÎncă nu există evaluări
- September 15, 2016Document20 paginiSeptember 15, 2016CassieÎncă nu există evaluări
- Combat Lifesaver Guide to One-Person Bag Valve Mask VentilationDocument1 paginăCombat Lifesaver Guide to One-Person Bag Valve Mask VentilationSae TumÎncă nu există evaluări
- Detyre Kursi: Lenda:AnglishtDocument11 paginiDetyre Kursi: Lenda:AnglishtFjorelo FeroÎncă nu există evaluări
- BHP Formula No 55 Refrence - Bioplasgen 20 Homeopathic Medicine For Skin DiseasesDocument2 paginiBHP Formula No 55 Refrence - Bioplasgen 20 Homeopathic Medicine For Skin DiseasesKazy Habibur RahmanÎncă nu există evaluări