S-ar putea să vă placă și
- Benign and Pathological Chromosomal Imbalances: Microscopic and Submicroscopic Copy Number Variations (CNVs) in Genetics and CounselingDe la EverandBenign and Pathological Chromosomal Imbalances: Microscopic and Submicroscopic Copy Number Variations (CNVs) in Genetics and CounselingÎncă nu există evaluări
- 10 2018 DiGeorge Molecular Genetics of 22q11.2 Deletion SyndromeDocument12 pagini10 2018 DiGeorge Molecular Genetics of 22q11.2 Deletion SyndromeTRÍ LÊ VĂNÎncă nu există evaluări
- Cat Eye SyndromeDocument3 paginiCat Eye SyndromeNajia ChoudhuryÎncă nu există evaluări
- Understanding Familial and Non-Familial Renal Cell CancerDocument10 paginiUnderstanding Familial and Non-Familial Renal Cell CancerVanroÎncă nu există evaluări
- Abstract BookDocument90 paginiAbstract BookPopescu RoxanaÎncă nu există evaluări
- 2013 Delgado MolSyndromolDocument5 pagini2013 Delgado MolSyndromolEAPÎncă nu există evaluări
- Citometria 2005Document8 paginiCitometria 2005Leslie AraujoÎncă nu există evaluări
- Detection of Deletions at 7q11.23 in Williams-BeurenDocument6 paginiDetection of Deletions at 7q11.23 in Williams-BeurenAmisadai ArévaloÎncă nu există evaluări
- WBSCR14, A Putative Transcription Factor GeneDocument8 paginiWBSCR14, A Putative Transcription Factor GeneRaymond LaBoyÎncă nu există evaluări
- Velo-Cardio-Facial Syndrome (22q11.2 Deletion Syndrome)Document24 paginiVelo-Cardio-Facial Syndrome (22q11.2 Deletion Syndrome)FantasyWorldÎncă nu există evaluări
- Objectives:: Clinical Relevance of MYC, BCL2 and BCL6 Rearrangements and Amplification in Diffuse Large B-Cell LymphomaDocument1 paginăObjectives:: Clinical Relevance of MYC, BCL2 and BCL6 Rearrangements and Amplification in Diffuse Large B-Cell LymphomaEddie CaptainÎncă nu există evaluări
- A Fifth Locus For Otosclerosis, OTSC5, Maps To Chromosome 3q22-24Document4 paginiA Fifth Locus For Otosclerosis, OTSC5, Maps To Chromosome 3q22-24Ita MaghfirahÎncă nu există evaluări
- Chronic Lymphocytic Leukemia: A Clinical and Molecular Heterogenous DiseaseDocument14 paginiChronic Lymphocytic Leukemia: A Clinical and Molecular Heterogenous DiseaseCallisthenisLeventisÎncă nu există evaluări
- Hum Mol Genet 2021 30 2215Document10 paginiHum Mol Genet 2021 30 2215Biblioteca FleniÎncă nu există evaluări
- Rodrguez Vicente 2013Document14 paginiRodrguez Vicente 2013Jose AbadiaÎncă nu există evaluări
- Anemia de Cells Falc en Urgencias Clínicas NADocument19 paginiAnemia de Cells Falc en Urgencias Clínicas NACarlos Andres Barrera ParraÎncă nu există evaluări
- Clinical Implications of The Genetic Architecture of Dilated CardiomyopathyDocument11 paginiClinical Implications of The Genetic Architecture of Dilated CardiomyopathyIsabella-Diana ChelbanÎncă nu există evaluări
- Research Article: Williams-Beuren Syndrome: A Clinical Study of 55 Brazilian Patients and The Diagnostic Use of MLPADocument7 paginiResearch Article: Williams-Beuren Syndrome: A Clinical Study of 55 Brazilian Patients and The Diagnostic Use of MLPApejavzÎncă nu există evaluări
- Minimal Residual Disease Detection of Myeloma Using Sequencing of Immunoglobulin Heavy Chain Gene VDJ RegionsDocument18 paginiMinimal Residual Disease Detection of Myeloma Using Sequencing of Immunoglobulin Heavy Chain Gene VDJ RegionsSeungman ParkÎncă nu există evaluări
- BMC Medical Genetics: Pathogenesis of Vestibular Schwannoma in Ring Chromosome 22Document7 paginiBMC Medical Genetics: Pathogenesis of Vestibular Schwannoma in Ring Chromosome 22Lulu SupergirlÎncă nu există evaluări
- Zeng 2016Document7 paginiZeng 2016Thanasis PapatheodorouÎncă nu există evaluări
- Neuronal Migration Genes and A Familial Translocation T (3 17) : Candidate Genes Implicated in The PhenotypeDocument11 paginiNeuronal Migration Genes and A Familial Translocation T (3 17) : Candidate Genes Implicated in The PhenotypeTiago ChavesÎncă nu există evaluări
- Medicine - IJGMP - Update On Arrhythmogenic Ventricular Dysplasia - Francesco Massoni - RomeDocument10 paginiMedicine - IJGMP - Update On Arrhythmogenic Ventricular Dysplasia - Francesco Massoni - Romeiaset123Încă nu există evaluări
- Zufferey. 2012.A 600 KB Deletion Syndrome at 16p11.2 Leads To Energy Imbalance and Neuropsychiatric DisordersDocument10 paginiZufferey. 2012.A 600 KB Deletion Syndrome at 16p11.2 Leads To Energy Imbalance and Neuropsychiatric DisordersEccoÎncă nu există evaluări
- B-Lymphoblastic Leukemia Presenting With An IsoderDocument5 paginiB-Lymphoblastic Leukemia Presenting With An Isoderzehraamber110Încă nu există evaluări
- 2016 Hussein MolCytogenetDocument13 pagini2016 Hussein MolCytogenetEduardo PasteneÎncă nu există evaluări
- Newborn Infant With Inherited Ring and de Novo Interstitial Deletion On Homologous Chromosome 22sDocument4 paginiNewborn Infant With Inherited Ring and de Novo Interstitial Deletion On Homologous Chromosome 22sAraceli Enríquez OvandoÎncă nu există evaluări
- Congenital Adrenal Hyperplasia Due To 21-Hydroxylase DeficiencyDocument14 paginiCongenital Adrenal Hyperplasia Due To 21-Hydroxylase DeficiencyveraÎncă nu există evaluări
- CSF3R 3Document10 paginiCSF3R 3AndikhaÎncă nu există evaluări
- Polycystic Kidney - Eight CasesDocument11 paginiPolycystic Kidney - Eight CasesKAINAT ANJUMÎncă nu există evaluări
- Pathogenesis and Management of Essential Thrombocythemia: Philip A. Beer and Anthony R. GreenDocument8 paginiPathogenesis and Management of Essential Thrombocythemia: Philip A. Beer and Anthony R. GreenHoopmen SilaenÎncă nu există evaluări
- Cah Nejm 2020Document14 paginiCah Nejm 2020Nor Azmi KamaruddinÎncă nu există evaluări
- Lab 1 (Article)Document5 paginiLab 1 (Article)Vignesh R.PuvanesuaranÎncă nu există evaluări
- Zayac Retrospective HGBL Blood Adv 2023Document14 paginiZayac Retrospective HGBL Blood Adv 2023Cabinet Dr ZdrengheaÎncă nu există evaluări
- 1 s2.0 S0006497119353972 MainDocument3 pagini1 s2.0 S0006497119353972 Mainmiltoncaballero5Încă nu există evaluări
- CRLF2 and IKZF1 Abnormalities in Mexican Children With AcuteDocument12 paginiCRLF2 and IKZF1 Abnormalities in Mexican Children With Acutemejia_jpÎncă nu există evaluări
- CML Case Study - 3rd GalleyDocument2 paginiCML Case Study - 3rd GalleycegfracÎncă nu există evaluări
- LinfomaDocument9 paginiLinfomaRocio HaroÎncă nu există evaluări
- NIH Public Access: Author ManuscriptDocument15 paginiNIH Public Access: Author ManuscriptJaclyn OlsenÎncă nu există evaluări
- DHH-RHEBL1 Fusion Transcript A Novel Recurrent AML Ricardo, 2013Document9 paginiDHH-RHEBL1 Fusion Transcript A Novel Recurrent AML Ricardo, 2013mrkhprojectÎncă nu există evaluări
- Blood 2021 Gemenetzi Et AlDocument10 paginiBlood 2021 Gemenetzi Et Almgounari7293Încă nu există evaluări
- Milno Et AlDocument9 paginiMilno Et AlLara SilveiraÎncă nu există evaluări
- Regular ArticleDocument11 paginiRegular ArticleOmar FernándezÎncă nu există evaluări
- Sarcoidosis CardíacaDocument14 paginiSarcoidosis Cardíacaluiserikolivares1390Încă nu există evaluări
- Two Atypical Cases of Classical Hairy Cell LeukaemDocument8 paginiTwo Atypical Cases of Classical Hairy Cell Leukaemwindusanjaya606Încă nu există evaluări
- Int J Dermatology - 2015 - Bonchak - Calciphylaxis A Case Series and The Role of Radiology in DiagnosisDocument5 paginiInt J Dermatology - 2015 - Bonchak - Calciphylaxis A Case Series and The Role of Radiology in DiagnosisRichard LiuÎncă nu există evaluări
- Abnormalities of RUNX1, CBFB, CEBPA, and NPM1 Genes in Acute Myeloid LeukemiaDocument10 paginiAbnormalities of RUNX1, CBFB, CEBPA, and NPM1 Genes in Acute Myeloid LeukemiaAndikhaÎncă nu există evaluări
- Cytogenetic Analysis in The Diagnosis of Acute Leukemia: Sverre Heim, FelixDocument9 paginiCytogenetic Analysis in The Diagnosis of Acute Leukemia: Sverre Heim, FelixEnas KharbotlyÎncă nu există evaluări
- Interrogation of Cancer Gene Dependencies Reveals Novel Paralog Interactions of Autosome and Sex Chromosome Encoded GenesDocument40 paginiInterrogation of Cancer Gene Dependencies Reveals Novel Paralog Interactions of Autosome and Sex Chromosome Encoded GenesMadhwesh CrÎncă nu există evaluări
- CRISPR-Cas9 Gene Editing for Sickle Cell Disease and β-ThalassemiaDocument9 paginiCRISPR-Cas9 Gene Editing for Sickle Cell Disease and β-ThalassemiadrsourabhsinghÎncă nu există evaluări
- Cloning Candidate Genes Involved in The Beckwith-Wiedemann Syndrome and Childhood TumorsDocument3 paginiCloning Candidate Genes Involved in The Beckwith-Wiedemann Syndrome and Childhood TumorsPaul AsturbiarisÎncă nu există evaluări
- Acute Myeloid Leukemia With Myelodysplasia RelatedDocument6 paginiAcute Myeloid Leukemia With Myelodysplasia RelatedAgus WiniÎncă nu există evaluări
- Echocardiographic Categorization of Morphology of Mitral Valve in Patients With Degenerative Mitral Alve Disease: Correlation With Clinical FeaturesDocument4 paginiEchocardiographic Categorization of Morphology of Mitral Valve in Patients With Degenerative Mitral Alve Disease: Correlation With Clinical FeaturesBIOMEDSCIDIRECT PUBLICATIONSÎncă nu există evaluări
- Anucleate Platelets Generate ProgenyDocument3 paginiAnucleate Platelets Generate ProgenyAlejandro BarretoÎncă nu există evaluări
- Shah Khan 2012Document8 paginiShah Khan 2012Roberson GuimaraesÎncă nu există evaluări
- Double Hit Lymphomas Clinical Morphological Immunohistochemical and Cytogenetic Study in A Series of Brazilian Patients With High Grade Non Hodgkin LymphomaDocument9 paginiDouble Hit Lymphomas Clinical Morphological Immunohistochemical and Cytogenetic Study in A Series of Brazilian Patients With High Grade Non Hodgkin LymphomaCierre WesleyÎncă nu există evaluări
- Revista Española de Cardiología (English Edition) : Volume 62, Issue 3Document3 paginiRevista Española de Cardiología (English Edition) : Volume 62, Issue 3Yave Aro RodriguezÎncă nu există evaluări
- Sindrome MielodisplasicosDocument15 paginiSindrome MielodisplasicosMatias FlammÎncă nu există evaluări
- CAH Kulll 2020 Review 10.1056@NEJMra1909786Document14 paginiCAH Kulll 2020 Review 10.1056@NEJMra1909786MCuk2606Încă nu există evaluări
- CAG Repeat Instability at SCA2 Locus - Anchoring CAA Interruptions and Linked Single Nucleotide PolymorphismsDocument10 paginiCAG Repeat Instability at SCA2 Locus - Anchoring CAA Interruptions and Linked Single Nucleotide PolymorphismsSarahÎncă nu există evaluări
- Ascitis en NiñosDocument7 paginiAscitis en NiñosBrenda CaraveoÎncă nu există evaluări
- Hypercholesterolemia Among Children: When Is It High, and When Is It Really High?Document4 paginiHypercholesterolemia Among Children: When Is It High, and When Is It Really High?Brenda CaraveoÎncă nu există evaluări
- Treatment of Allergic Rhinitis PDFDocument23 paginiTreatment of Allergic Rhinitis PDFLydia MamurÎncă nu există evaluări
- Abdominal Surgical Emergencies in Infants and Young ChildrenDocument27 paginiAbdominal Surgical Emergencies in Infants and Young ChildrenBrenda CaraveoÎncă nu există evaluări
- Adenomegalias 08Document10 paginiAdenomegalias 08Brenda CaraveoÎncă nu există evaluări
- Alt Coagulacion 08Document12 paginiAlt Coagulacion 08Brenda CaraveoÎncă nu există evaluări
- Clinical Implications of Cross Reactive Food AllergensDocument10 paginiClinical Implications of Cross Reactive Food AllergensBrenda CaraveoÎncă nu există evaluări
- FWD Medical Rider enDocument12 paginiFWD Medical Rider enKhairul HMSBÎncă nu există evaluări
- Caffein Intox PDFDocument3 paginiCaffein Intox PDFSejahtera SurbaktiÎncă nu există evaluări
- CHN Practice Exam 1Document7 paginiCHN Practice Exam 1Domeyeg, Clyde Mae W.100% (1)
- Rheumatic Fever PDFDocument3 paginiRheumatic Fever PDFDoc JT BylnÎncă nu există evaluări
- Silkworm in NepalDocument11 paginiSilkworm in NepalRitesh Raj Shrestha50% (4)
- Chapter 12 Glencoe Biology Study GuideDocument11 paginiChapter 12 Glencoe Biology Study GuideabdulazizalobaidiÎncă nu există evaluări
- Final Coaching NP4 Set 1Document13 paginiFinal Coaching NP4 Set 1STEFFI GABRIELLE GOLEZÎncă nu există evaluări
- Master The Perfec Tnight of SleepDocument10 paginiMaster The Perfec Tnight of SleepPaulo André Prada de CamargoÎncă nu există evaluări
- 2018 2019 Daily Lesson Log AutosavedDocument322 pagini2018 2019 Daily Lesson Log AutosavedAinah 16Încă nu există evaluări
- Pathomorphology MCQSDocument191 paginiPathomorphology MCQSNana BentilÎncă nu există evaluări
- UntitledDocument44 paginiUntitled제올Încă nu există evaluări
- Mci Mbbs SyllabusDocument110 paginiMci Mbbs Syllabusmetabolicmedicine100% (1)
- Scid 5 CVDocument26 paginiScid 5 CVZubair Mahmood KamalÎncă nu există evaluări
- PHD Research ProposalDocument8 paginiPHD Research ProposalMalik Ijaz Khan100% (1)
- Full Download Test Bank Health Assessment For Nursing Practice 6th Edition Wilson PDF Full ChapterDocument36 paginiFull Download Test Bank Health Assessment For Nursing Practice 6th Edition Wilson PDF Full Chapterbeatermany.imubd2100% (19)
- Ghidul de Prevenire A Miopiei - de Dr. Kaisu VIIKARI - Cu Coperta in RomanaDocument104 paginiGhidul de Prevenire A Miopiei - de Dr. Kaisu VIIKARI - Cu Coperta in Romanadlucian82Încă nu există evaluări
- Endocrine Glands PDFDocument99 paginiEndocrine Glands PDFXochitl ZambranoÎncă nu există evaluări
- Mekong TMBO 02 Maret 2021Document14 paginiMekong TMBO 02 Maret 2021Yolanda RahayuÎncă nu există evaluări
- Deeper Trial - Asco 2021Document26 paginiDeeper Trial - Asco 2021Vuong Dinh Thy HaoÎncă nu există evaluări
- Normal Periodontium 1Document49 paginiNormal Periodontium 1mdio midoÎncă nu există evaluări
- Case Study Presentation-Mindy Duran-FinalDocument30 paginiCase Study Presentation-Mindy Duran-Finalapi-278622211Încă nu există evaluări
- Spring2013 - Wise Traditions PDFDocument128 paginiSpring2013 - Wise Traditions PDFfumiÎncă nu există evaluări
- Devil You Know PDFDocument269 paginiDevil You Know PDFElissaÎncă nu există evaluări
- Cipriani Et Al-2013-Cochrane Database of Systematic ReviewsDocument52 paginiCipriani Et Al-2013-Cochrane Database of Systematic ReviewsfiskaderishaÎncă nu există evaluări
- Bronchiectasis: Citadel B. Rabanes, MD First Year JRRMMC-Department of RadiologyDocument34 paginiBronchiectasis: Citadel B. Rabanes, MD First Year JRRMMC-Department of Radiologyadhik_deepak100% (1)
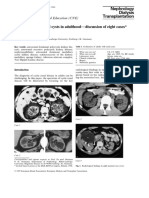
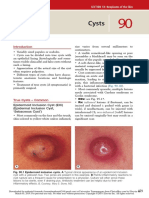
- CystsDocument5 paginiCystsranindita maulyaÎncă nu există evaluări
- Block 4Document106 paginiBlock 4Dr. Megha RichhariyaÎncă nu există evaluări
- Refers To Leisure Pursuits Engaged in The Outdoors, Often in Natural or Semi-Natural Settings Out of TownDocument21 paginiRefers To Leisure Pursuits Engaged in The Outdoors, Often in Natural or Semi-Natural Settings Out of TownSam CajesÎncă nu există evaluări
- Ultimate Guide To Surviving in The WildDocument175 paginiUltimate Guide To Surviving in The WildSal Ot100% (1)
- Pharmacovigilance - An Overview: Dr. Sukanta Sen MD, DNB, DM (Clin Pharmacology)Document31 paginiPharmacovigilance - An Overview: Dr. Sukanta Sen MD, DNB, DM (Clin Pharmacology)FranchescaAmoréBrodithe100% (1)