S-ar putea să vă placă și
- Genetic Variation in Taste and Its Influence On Food SelectionDocument13 paginiGenetic Variation in Taste and Its Influence On Food Selectionmarine2006Încă nu există evaluări
- Water Taste: The Importance of Osmotic Sensing in The Oral CavityDocument6 paginiWater Taste: The Importance of Osmotic Sensing in The Oral CavityDwina RahmawatiÎncă nu există evaluări
- Making Sense of TasteDocument10 paginiMaking Sense of TastetufekciosmanÎncă nu există evaluări
- Genetics of PerceptionDocument6 paginiGenetics of PerceptiondionysiaÎncă nu există evaluări
- Fla Via 3Document7 paginiFla Via 3adeflavÎncă nu există evaluări
- TASRs - Gustatory and Extragustatory Functions#Document10 paginiTASRs - Gustatory and Extragustatory Functions#Luciana ParadelaÎncă nu există evaluări
- Unit-8 - Lung and Volume CapacityDocument14 paginiUnit-8 - Lung and Volume CapacityQ O R Y A N IÎncă nu există evaluări
- The Physiology of Taste in Fish Potential Implications For Feeding Stimulation and Gut Chemical SensingDocument18 paginiThe Physiology of Taste in Fish Potential Implications For Feeding Stimulation and Gut Chemical SensingDewi NurhayatiÎncă nu există evaluări
- Taste & FlavourDocument10 paginiTaste & Flavourjcoppala4476Încă nu există evaluări
- The Role of Saliva in Food Sensory Perception: Relevant Knowledge To Design Healthy FoodsDocument9 paginiThe Role of Saliva in Food Sensory Perception: Relevant Knowledge To Design Healthy FoodsTanChantreaÎncă nu există evaluări
- Taste: Topics in Chemical Biology: Advanced ArticleDocument12 paginiTaste: Topics in Chemical Biology: Advanced ArticleqhqhqÎncă nu există evaluări
- Presynaptic (Type III) Cells in Mouse Taste Buds Sense Sour (Acid) TasteDocument10 paginiPresynaptic (Type III) Cells in Mouse Taste Buds Sense Sour (Acid) TasteSusana AdameÎncă nu există evaluări
- Klaus Reutter-Fish Chemosenses (Teleostean Fish Biology A Comprehensive Examination of Major Taxa) (2005)Document357 paginiKlaus Reutter-Fish Chemosenses (Teleostean Fish Biology A Comprehensive Examination of Major Taxa) (2005)Beatrice SavaÎncă nu există evaluări
- Accepted Manuscript: Food ChemistryDocument21 paginiAccepted Manuscript: Food ChemistryAnne Sophie ArcigaÎncă nu există evaluări
- Accepted Manuscript: Food ChemistryDocument21 paginiAccepted Manuscript: Food ChemistryAnne Sophie ArcigaÎncă nu există evaluări
- Accepted Manuscript: Food ChemistryDocument21 paginiAccepted Manuscript: Food ChemistryAnne Sophie ArcigaÎncă nu există evaluări
- ID Leksikal Taksonomi Makanan Dalam BahasaDocument15 paginiID Leksikal Taksonomi Makanan Dalam BahasaEkklesia UmbohÎncă nu există evaluări
- Salt Sensation and RegulationDocument12 paginiSalt Sensation and RegulationВладимир ДружининÎncă nu există evaluări
- Menis-Henrique2020 Flavor PDFDocument5 paginiMenis-Henrique2020 Flavor PDFRizha Gustian FirdausÎncă nu există evaluări
- DysgeusiaDocument14 paginiDysgeusiasandaÎncă nu există evaluări
- The Science of TasteDocument107 paginiThe Science of TasteAnonymous U9aZZmeNrEÎncă nu există evaluări
- Coffee Roasting and GrindingDocument182 paginiCoffee Roasting and GrindingTsegaye MulugetaÎncă nu există evaluări
- Food PairingDocument5 paginiFood PairingScribdTranslationsÎncă nu există evaluări
- Foods: Food Odours Direct Specific AppetiteDocument10 paginiFoods: Food Odours Direct Specific Appetitekucing menariÎncă nu există evaluări
- Confusing Tastes With Flavours PDFDocument39 paginiConfusing Tastes With Flavours PDFhattyuuÎncă nu există evaluări
- Thesis Robin FondbergDocument67 paginiThesis Robin FondbergJOHANNAH MARIE TOMASÎncă nu există evaluări
- Maheswaran2014 PDFDocument5 paginiMaheswaran2014 PDFMaqbul AlamÎncă nu există evaluări
- Science Research Chapter 3-2-11Document16 paginiScience Research Chapter 3-2-11fffaccurateÎncă nu există evaluări
- Todd 2012Document4 paginiTodd 2012Spoti PremiÎncă nu există evaluări
- Stomach Tastes 2020Document4 paginiStomach Tastes 2020Владимир ДружининÎncă nu există evaluări
- Conditioned Taste Aversions: SciencedirectDocument9 paginiConditioned Taste Aversions: SciencedirectBotez MartaÎncă nu există evaluări
- Biochem Week 1Document17 paginiBiochem Week 1divineÎncă nu există evaluări
- ProbioticoDocument15 paginiProbioticoJose Vicente Benitez MoralesÎncă nu există evaluări
- On Digestive Proteolysis: Being the Cartwright Lectures for 1894De la EverandOn Digestive Proteolysis: Being the Cartwright Lectures for 1894Încă nu există evaluări
- Miracle Fruit ResearchDocument9 paginiMiracle Fruit ResearchKyla de SilvaÎncă nu există evaluări
- Keywords: Taste, Gustatory, Water, Nucleus of The Solitary Tract, Parabrachial Nucleus of TheDocument14 paginiKeywords: Taste, Gustatory, Water, Nucleus of The Solitary Tract, Parabrachial Nucleus of ThekikÎncă nu există evaluări
- Flavor Network and The Principles of Food Pairing: Scientific ReportsDocument7 paginiFlavor Network and The Principles of Food Pairing: Scientific ReportsAnonymous oIa2cFuoÎncă nu există evaluări
- Analysis of Volatile Organic Compounds in Human Saliva by A Static Sorptive Extraction Method and Gas Chromatography-Mass SpectrometryDocument8 paginiAnalysis of Volatile Organic Compounds in Human Saliva by A Static Sorptive Extraction Method and Gas Chromatography-Mass SpectrometryAndré Felippe XavierÎncă nu există evaluări
- Chap1 IntroductionDocument5 paginiChap1 Introductionsyg0302Încă nu există evaluări
- Paleolithic-Nutrition-Twenty-Five Years-Later-Eaton PDFDocument9 paginiPaleolithic-Nutrition-Twenty-Five Years-Later-Eaton PDFFran HrzicÎncă nu există evaluări
- Metabolic Inhibitors V1: A Comprehensive TreatiseDe la EverandMetabolic Inhibitors V1: A Comprehensive TreatiseR HochsterÎncă nu există evaluări
- Molecular Gastronomy Used in BakeryDocument64 paginiMolecular Gastronomy Used in BakeryIyer Guhan67% (3)
- Journal of Fish Biology - 2019 - Kasumyan - The Taste System in Fishes and The Effects of Environmental VariablesDocument24 paginiJournal of Fish Biology - 2019 - Kasumyan - The Taste System in Fishes and The Effects of Environmental VariablesDewi NurhayatiÎncă nu există evaluări
- The Multisensory Perception of Flavour: Charles Spence On His Mouth-Watering ResearchDocument4 paginiThe Multisensory Perception of Flavour: Charles Spence On His Mouth-Watering Researchapi-109637900Încă nu există evaluări
- Bile Acids Review 2009-1Document13 paginiBile Acids Review 2009-1cs1900Încă nu există evaluări
- AirVeda: Ancient & New Medical Wisdom, Digestion & Gas, Volume OneDe la EverandAirVeda: Ancient & New Medical Wisdom, Digestion & Gas, Volume OneÎncă nu există evaluări
- Reflections On Nutrition in Biological Anthropology.Document10 paginiReflections On Nutrition in Biological Anthropology.José OrtízÎncă nu există evaluări
- 1675 FullDocument14 pagini1675 FullPallavi DubeyÎncă nu există evaluări
- Malika-AThe Multisensory Perception of Flavoruvray-Auvray Spence 2008 CCDocument16 paginiMalika-AThe Multisensory Perception of Flavoruvray-Auvray Spence 2008 CCvoodoodaveÎncă nu există evaluări
- European Journal of Pharmaceutical SciencesDocument8 paginiEuropean Journal of Pharmaceutical SciencesrezqÎncă nu există evaluări
- Molecular Gastronomy Used in BakeryDocument64 paginiMolecular Gastronomy Used in Bakerysmyrna_exodus100% (1)
- Food Sciences and Nutrition. 2003 881-887.: Daftar PustakaDocument2 paginiFood Sciences and Nutrition. 2003 881-887.: Daftar PustakaStephanie Gabriella KamÎncă nu există evaluări
- AirVeda: Ancient & New Medical Wisdom, Digestion & Gas, Volume TwoDe la EverandAirVeda: Ancient & New Medical Wisdom, Digestion & Gas, Volume TwoÎncă nu există evaluări
- Age-Related Deficits in Taste and SmellDocument11 paginiAge-Related Deficits in Taste and SmellMARA ROMEROÎncă nu există evaluări
- Nutrition and Taste and Smell Dysfunction: SciencedirectDocument8 paginiNutrition and Taste and Smell Dysfunction: SciencedirectEster HarsiwiÎncă nu există evaluări
- Am J Physiol Regul Integr Comp Physiol-2001-Kristan-R502-10Document10 paginiAm J Physiol Regul Integr Comp Physiol-2001-Kristan-R502-10EstuardoAÎncă nu există evaluări
- Right Food For A Pregnant Woman According To AyurvedaDocument5 paginiRight Food For A Pregnant Woman According To AyurvedaJaime BallesterosÎncă nu există evaluări
- Dayal Bagh: Spircon 2013: Book of AbstractsDocument202 paginiDayal Bagh: Spircon 2013: Book of AbstractsSant Mat100% (2)
- Natural Ways To Heal Pains According To AyurvedaDocument8 paginiNatural Ways To Heal Pains According To AyurvedaJaime BallesterosÎncă nu există evaluări
- Yoga, Ayurveda and Exercise Are The Basis of Good HealthDocument10 paginiYoga, Ayurveda and Exercise Are The Basis of Good Healthvijender AtriÎncă nu există evaluări
- Rejuvenating Mind, Body and Soul in Old Age Through Yoga and AyurvedaDocument3 paginiRejuvenating Mind, Body and Soul in Old Age Through Yoga and AyurvedaJaime BallesterosÎncă nu există evaluări
- Adaptógenos Plantas Medicinales Tradicionales PDFDocument14 paginiAdaptógenos Plantas Medicinales Tradicionales PDFJaime BallesterosÎncă nu există evaluări
- Health Benefits of Pippali Fruit According To AyurvedaDocument9 paginiHealth Benefits of Pippali Fruit According To AyurvedaJaime BallesterosÎncă nu există evaluări
- How Shaolin Kung Fu Came From IndiaDocument6 paginiHow Shaolin Kung Fu Came From IndiaRicardo IglesiasÎncă nu există evaluări
- Harvard Neuroscientist Meditation and The BrainDocument7 paginiHarvard Neuroscientist Meditation and The BrainClaudio Javier FredesÎncă nu există evaluări
- Hindu Scriptures Mention Gravity 1Document10 paginiHindu Scriptures Mention Gravity 1Jaime BallesterosÎncă nu există evaluări
- How The Sense of Taste Has Shaped Who We AreDocument6 paginiHow The Sense of Taste Has Shaped Who We AreJaime BallesterosÎncă nu există evaluări
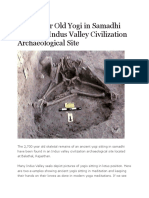
- 2,700 Year Old Yogi in Samadhi Found in Indus ValleyDocument12 pagini2,700 Year Old Yogi in Samadhi Found in Indus ValleyMisterÎncă nu există evaluări
- Potent Health Benefits of Six Traditional Ayurvedic HerbsDocument4 paginiPotent Health Benefits of Six Traditional Ayurvedic HerbsJaime Ballesteros100% (1)
- Health Benefits of Wheat and Its Nutritional ValueDocument13 paginiHealth Benefits of Wheat and Its Nutritional ValueJaime BallesterosÎncă nu există evaluări
- The Antiquity of Vedic CivilizationDocument9 paginiThe Antiquity of Vedic CivilizationJaime BallesterosÎncă nu există evaluări
- Four Ayurvedic Herbs To Enhance Brain Power NaturallyDocument6 paginiFour Ayurvedic Herbs To Enhance Brain Power NaturallyJaime BallesterosÎncă nu există evaluări
- Health and Exercise According To Ayurveda and Yoga ShastraDocument3 paginiHealth and Exercise According To Ayurveda and Yoga ShastraJaime BallesterosÎncă nu există evaluări
- Arjuna Tree Qualities, Benefits and Natural Remedies According To AyurvedaDocument8 paginiArjuna Tree Qualities, Benefits and Natural Remedies According To AyurvedaJaime BallesterosÎncă nu există evaluări
- Benefits of Cow Protection As Per ScienceDocument10 paginiBenefits of Cow Protection As Per ScienceJaime BallesterosÎncă nu există evaluări
- Benefits of Eating Basmati Rice According To AyurvedaDocument4 paginiBenefits of Eating Basmati Rice According To AyurvedaJaime BallesterosÎncă nu există evaluări
- HalasanaDocument6 paginiHalasanaJaime BallesterosÎncă nu există evaluări
- HalasanaDocument6 paginiHalasanaJaime BallesterosÎncă nu există evaluări
- The Vedic ShakhasDocument7 paginiThe Vedic ShakhasJaime Ballesteros100% (1)
- Benefits of Ghee According To AyurvedaDocument9 paginiBenefits of Ghee According To AyurvedaJaime BallesterosÎncă nu există evaluări
- Emergent Phenomena in Mesoscopic SystemsDocument43 paginiEmergent Phenomena in Mesoscopic SystemsJaime BallesterosÎncă nu există evaluări
- Stephen Hawking - There Are No Black HolesDocument4 paginiStephen Hawking - There Are No Black HolesJaime BallesterosÎncă nu există evaluări
- How Your Brain Learns PhysicsDocument2 paginiHow Your Brain Learns PhysicsJaime Ballesteros100% (1)
- Non-Newtonian Fluid PDFDocument6 paginiNon-Newtonian Fluid PDFJaime BallesterosÎncă nu există evaluări
- Bacteriofagos Adheridos A Celulas de Mucosa Intestinal InmunologiaDocument6 paginiBacteriofagos Adheridos A Celulas de Mucosa Intestinal InmunologiaJaime BallesterosÎncă nu există evaluări
- Chapter 5 - CheerdanceDocument10 paginiChapter 5 - CheerdanceJoana CampoÎncă nu există evaluări
- CUBE Dealer Book 2009Document280 paginiCUBE Dealer Book 2009maikruetzÎncă nu există evaluări
- Crisis of The World Split Apart: Solzhenitsyn On The WestDocument52 paginiCrisis of The World Split Apart: Solzhenitsyn On The WestdodnkaÎncă nu există evaluări
- Art of War Day TradingDocument17 paginiArt of War Day TradingChrispen MoyoÎncă nu există evaluări
- Roles and Responsibilities of An InstructorDocument4 paginiRoles and Responsibilities of An InstructorMohanlal SainiÎncă nu există evaluări
- Ismb ItpDocument3 paginiIsmb ItpKumar AbhishekÎncă nu există evaluări
- Stonehell Dungeon 1 Down Night Haunted Halls (LL)Document138 paginiStonehell Dungeon 1 Down Night Haunted Halls (LL)some dude100% (9)
- Flow of FoodDocument2 paginiFlow of FoodGenevaÎncă nu există evaluări
- Presenters: Horace M. Estrella Jay Mart A. Lazana Princess Camille R. HipolitoDocument23 paginiPresenters: Horace M. Estrella Jay Mart A. Lazana Princess Camille R. HipolitoHorace EstrellaÎncă nu există evaluări
- Notice: Premerger Notification Waiting Periods Early TerminationsDocument5 paginiNotice: Premerger Notification Waiting Periods Early TerminationsJustia.comÎncă nu există evaluări
- FINAL BÁO-CÁO-THỰC-TẬP.editedDocument38 paginiFINAL BÁO-CÁO-THỰC-TẬP.editedngocthaongothi4Încă nu există evaluări
- Contemporary Philippine Arts From The Regions: Quarter 1Document11 paginiContemporary Philippine Arts From The Regions: Quarter 1JUN GERONAÎncă nu există evaluări
- (Isaac Asimov) How Did We Find Out About AntarcticDocument24 pagini(Isaac Asimov) How Did We Find Out About AntarcticDrBabu PSÎncă nu există evaluări
- LPS 1131-Issue 1.2-Requirements and Testing Methods For Pumps For Automatic Sprinkler Installation Pump Sets PDFDocument19 paginiLPS 1131-Issue 1.2-Requirements and Testing Methods For Pumps For Automatic Sprinkler Installation Pump Sets PDFHazem HabibÎncă nu există evaluări
- UAV Design TrainingDocument17 paginiUAV Design TrainingPritam AshutoshÎncă nu există evaluări
- Music 9 Q3 Mod4 Musical Elements of Given Romantic Period PiecesDocument19 paginiMusic 9 Q3 Mod4 Musical Elements of Given Romantic Period PiecesFinn Daniel Omayao100% (1)
- Adaptive Leadership: Leadership: Theory and PracticeDocument14 paginiAdaptive Leadership: Leadership: Theory and PracticeJose Daniel Quintero100% (1)
- Days Papers 2001Document341 paginiDays Papers 2001jorgefeitoza_hotmailÎncă nu există evaluări
- Barrett Beyond Psychometrics 2003 AugmentedDocument34 paginiBarrett Beyond Psychometrics 2003 AugmentedRoy Umaña CarrilloÎncă nu există evaluări
- A Teachers' Journey: Phenomenological Study On The Puritive Behavioral Standards of Students With Broken FamilyDocument11 paginiA Teachers' Journey: Phenomenological Study On The Puritive Behavioral Standards of Students With Broken FamilyNova Ariston100% (2)
- Directorate of Technical Education, Admission Committee For Professional Courses (ACPC), GujaratDocument2 paginiDirectorate of Technical Education, Admission Committee For Professional Courses (ACPC), GujaratgamailkabaaaapÎncă nu există evaluări
- Prognostic Factors and Management of Patients With Choanal AtresiaDocument7 paginiPrognostic Factors and Management of Patients With Choanal Atresiafarah maulida martaÎncă nu există evaluări
- Fake News Infographics by SlidesgoDocument33 paginiFake News Infographics by SlidesgoluanavicunhaÎncă nu există evaluări
- Reflection On The PoorDocument5 paginiReflection On The Poorapi-347831792Încă nu există evaluări
- Strategic Marketing: The University of Lahore (Islamabad Campus)Document3 paginiStrategic Marketing: The University of Lahore (Islamabad Campus)Sumaira IrshadÎncă nu există evaluări
- 1.co - Deb4113 - Industrial ManagementDocument10 pagini1.co - Deb4113 - Industrial ManagementrohaizadÎncă nu există evaluări
- Table of Specification 1st QDocument5 paginiTable of Specification 1st QVIRGILIO JR FABIÎncă nu există evaluări
- European Construction Sector Observatory: Country Profile MaltaDocument40 paginiEuropean Construction Sector Observatory: Country Profile MaltaRainbootÎncă nu există evaluări
- SecurityFund PPT 1.1Document13 paginiSecurityFund PPT 1.1Fmunoz MunozÎncă nu există evaluări
- BSH 7005-15Document129 paginiBSH 7005-15Mark InnesÎncă nu există evaluări