S-ar putea să vă placă și
- Reaction of ATCC 15697 To Oxidative Stress: Bifidobacterium Longum Subsp. Infantis StrainDocument15 paginiReaction of ATCC 15697 To Oxidative Stress: Bifidobacterium Longum Subsp. Infantis StrainMohammed SherifÎncă nu există evaluări
- GeLC-MS-based Proteomics of Chromobacterium Violaceum Comparison of Proteome Changes Elicited by Hydrogen PeroxidDocument12 paginiGeLC-MS-based Proteomics of Chromobacterium Violaceum Comparison of Proteome Changes Elicited by Hydrogen PeroxidLeidaiany SantosÎncă nu există evaluări
- 10 1016@j Ijbiomac 2020 05 135Document33 pagini10 1016@j Ijbiomac 2020 05 135MariÎncă nu există evaluări
- Zii 3122Document10 paginiZii 3122Muhammad Ricky RamadhianÎncă nu există evaluări
- LECTURA 1. Nicolaidis et al 2013. Resistance to antibiotics targeted to the bacterial cell wallDocument17 paginiLECTURA 1. Nicolaidis et al 2013. Resistance to antibiotics targeted to the bacterial cell wallAlmendra MosqueraÎncă nu există evaluări
- Study PlanDocument29 paginiStudy PlanMuhammad QasimÎncă nu există evaluări
- Colloids and Surfaces B: Biointerfaces: SciencedirectDocument9 paginiColloids and Surfaces B: Biointerfaces: SciencedirectMark AñazcoÎncă nu există evaluări
- Journal of Bacteriology-1983-Darveau-831.fullDocument8 paginiJournal of Bacteriology-1983-Darveau-831.fullanggi marlianaÎncă nu există evaluări
- Pharmaceuticals: Antimicrobial Peptides in 2014Document28 paginiPharmaceuticals: Antimicrobial Peptides in 2014Hermeson LimaÎncă nu există evaluări
- Taxonomy and Important Features of Probiotic Microorganisms in Food and NutritionDocument9 paginiTaxonomy and Important Features of Probiotic Microorganisms in Food and NutritionDhayu Mart Hindrasyah PandiaÎncă nu există evaluări
- EScholarship UC Item 372400z3Document24 paginiEScholarship UC Item 372400z3deslyati putri br sitepuÎncă nu există evaluări
- Mitochondrial Complex 1is Important For Plant Tolerance To Fungal Biotic StressDocument11 paginiMitochondrial Complex 1is Important For Plant Tolerance To Fungal Biotic StressSryahwa PublicationsÎncă nu există evaluări
- MC3 Group 5 Quiz #2Document6 paginiMC3 Group 5 Quiz #2Earl BenedictÎncă nu există evaluări
- Antioxidant Properties and Redox-Modulating Activity of ChitosanDocument9 paginiAntioxidant Properties and Redox-Modulating Activity of ChitosanGustavo RuizÎncă nu există evaluări
- Bioorganic & Medicinal Chemistry Letters: ArticleinfoDocument8 paginiBioorganic & Medicinal Chemistry Letters: ArticleinfoMehari AsratÎncă nu există evaluări
- Polya MineDocument10 paginiPolya MineLaili AlfinaÎncă nu există evaluări
- TMP 9 E27Document16 paginiTMP 9 E27FrontiersÎncă nu există evaluări
- CVBLIMDocument8 paginiCVBLIMfialophora0210Încă nu există evaluări
- A Culture-Independent Approach To Unravel Uncultured Bacteria and Functional Genes in A Complex Microbial CommunityDocument11 paginiA Culture-Independent Approach To Unravel Uncultured Bacteria and Functional Genes in A Complex Microbial CommunityBekele OljiraÎncă nu există evaluări
- Hylin DNA BBreportsDocument12 paginiHylin DNA BBreportsRamonquimicoÎncă nu există evaluări
- Metabolismo e MicrobiotaDocument14 paginiMetabolismo e MicrobiotaLívia Cristina Lima Dos SantosÎncă nu există evaluări
- Antibiotic Resistance Mechanisms in Bacteria: Biochemical and Genetic AspectsDocument11 paginiAntibiotic Resistance Mechanisms in Bacteria: Biochemical and Genetic AspectsD Wisam NajmÎncă nu există evaluări
- 2011 The LuxS-Dependent Quorum Sensing System Regulates Early Biofilm Formation by Streptococcus Pneumoniae Strain D39Document42 pagini2011 The LuxS-Dependent Quorum Sensing System Regulates Early Biofilm Formation by Streptococcus Pneumoniae Strain D39hazyaÎncă nu există evaluări
- Bioconversion of anthocyanins by probioticsDocument9 paginiBioconversion of anthocyanins by probioticsAnastasya AmandaÎncă nu există evaluări
- Activitty 1 LaboratoryDocument12 paginiActivitty 1 LaboratoryDarwin CauilanÎncă nu există evaluări
- Bioorganic & Medicinal ChemistryDocument6 paginiBioorganic & Medicinal ChemistryRuthaiwan KongcharoenÎncă nu există evaluări
- Planktocyclin, A Cyclooctapeptide Protease Inhibitor Produced by The Freshwater Cyanobacterium Planktothrix RubescensDocument5 paginiPlanktocyclin, A Cyclooctapeptide Protease Inhibitor Produced by The Freshwater Cyanobacterium Planktothrix RubescensJordana KalineÎncă nu există evaluări
- Full download book Recent Trends In Carbohydrate Chemistry Synthesis And Biomedical Applications Of Glycans And Glycoconjugates Pdf pdfDocument41 paginiFull download book Recent Trends In Carbohydrate Chemistry Synthesis And Biomedical Applications Of Glycans And Glycoconjugates Pdf pdfgeorge.berger158100% (18)
- Research Paper Pseudomonas AeruginosaDocument5 paginiResearch Paper Pseudomonas Aeruginosagatewivojez3100% (1)
- Production and Purification of Bacillus anthracis Protective AntigenDocument9 paginiProduction and Purification of Bacillus anthracis Protective AntigendyahTechÎncă nu există evaluări
- Caima2022 - Estabilidad y Actividad Biológica Mejoradas de Bacterioruberina en Nanovesículas MacrofagosDocument11 paginiCaima2022 - Estabilidad y Actividad Biológica Mejoradas de Bacterioruberina en Nanovesículas MacrofagosMark AñazcoÎncă nu există evaluări
- 1 s2.0 S0009308422000020 MainDocument10 pagini1 s2.0 S0009308422000020 MainHanan AqoubÎncă nu există evaluări
- 09campylobacter IDocument69 pagini09campylobacter Ianon_914901469Încă nu există evaluări
- Transcriptomic Analysis of The Under Oxidative Stress: Levilactobacillus Brevis 47f StrainDocument9 paginiTranscriptomic Analysis of The Under Oxidative Stress: Levilactobacillus Brevis 47f StrainMohammed SherifÎncă nu există evaluări
- Bacillus Subtillis PHDocument10 paginiBacillus Subtillis PHanon_816002920Încă nu există evaluări
- Bacterio Fag OsDocument11 paginiBacterio Fag OsTatiana CarrilloÎncă nu există evaluări
- Novel Biodemulsifier of Bacillus Mojavensis XH1 - Oxalate Decarboxylase With The Potential For Demulsification of Oilfield EmulsionDocument59 paginiNovel Biodemulsifier of Bacillus Mojavensis XH1 - Oxalate Decarboxylase With The Potential For Demulsification of Oilfield EmulsionMohamed AlhayaniÎncă nu există evaluări
- Bacterial LipasesDocument35 paginiBacterial LipasesLaura-Mihaela ChisÎncă nu există evaluări
- 2015-Appl Environ MicrobiolDocument9 pagini2015-Appl Environ MicrobiolMiguel Fernandez NiñoÎncă nu există evaluări
- Structural and Biological Aspects of Carotenoid CleavageDocument13 paginiStructural and Biological Aspects of Carotenoid CleavageMinh Chau NguyenÎncă nu există evaluări
- Biofilm Rpos PeroxidoDocument8 paginiBiofilm Rpos PeroxidoOmar MoralesÎncă nu există evaluări
- The Antioxidant Response of The Liver of Male SwisDocument7 paginiThe Antioxidant Response of The Liver of Male SwisLara Ayu LestariÎncă nu există evaluări
- Anaerobic Bacteriology: Clinical and Laboratory PracticeDe la EverandAnaerobic Bacteriology: Clinical and Laboratory PracticeÎncă nu există evaluări
- Oxidative Metabolism CanovaDocument13 paginiOxidative Metabolism CanovaA Sreenivasa ReddyÎncă nu există evaluări
- Aurantimonadaceae-Phylogenomics and Signature Proteins For The Alpha ProteobacteriaDocument20 paginiAurantimonadaceae-Phylogenomics and Signature Proteins For The Alpha ProteobacteriaAndrea EscobarÎncă nu există evaluări
- Article 1 PDFDocument19 paginiArticle 1 PDFAnonymous 5bgTK7qzsÎncă nu există evaluări
- Artigo 3Document6 paginiArtigo 3Uiara MariaÎncă nu există evaluări
- Cmls 2001 58 1650 65Document17 paginiCmls 2001 58 1650 65Nadia khalizaÎncă nu există evaluări
- Review: From Natural Product To Marketed Drug: The Tiacumicin OdysseyDocument14 paginiReview: From Natural Product To Marketed Drug: The Tiacumicin OdysseyJosué VelázquezÎncă nu există evaluări
- PQ 0403001990Document6 paginiPQ 0403001990Yuwono WibowoÎncă nu există evaluări
- Du Preez Et Al., 2008Document12 paginiDu Preez Et Al., 2008Linda ZommereÎncă nu există evaluări
- Campylobacter Jejuni Biofilms Up-Regulated in The Absence of The Stringent Response Utilize A Calcofluor White-Reactive PolysaccharideDocument11 paginiCampylobacter Jejuni Biofilms Up-Regulated in The Absence of The Stringent Response Utilize A Calcofluor White-Reactive PolysaccharideMichaelÎncă nu există evaluări
- Periodontology 2000 - 2022 - Pussinen - Periodontitis and Cardiometabolic Disorders The Role of Lipopolysaccharide andDocument22 paginiPeriodontology 2000 - 2022 - Pussinen - Periodontitis and Cardiometabolic Disorders The Role of Lipopolysaccharide andDr.Reshma RajendranÎncă nu există evaluări
- Said AhmedDocument19 paginiSaid Ahmedahmed mahdiÎncă nu există evaluări
- Metabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsDe la EverandMetabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsEndre A. BalazsÎncă nu există evaluări
- Fluid Mechanics, Bernoulli's Principle and Equation of ContinuityDocument10 paginiFluid Mechanics, Bernoulli's Principle and Equation of ContinuitySheerin SulthanaÎncă nu există evaluări
- J. Gen. Appl. Microbiol., 15, 387-398 (1969)Document12 paginiJ. Gen. Appl. Microbiol., 15, 387-398 (1969)Sheerin SulthanaÎncă nu există evaluări
- Trade-Related Intellectual Property Rights (Trips)Document20 paginiTrade-Related Intellectual Property Rights (Trips)Sheerin SulthanaÎncă nu există evaluări
- 32721Document33 pagini32721prasadbheemÎncă nu există evaluări
- Appl. Microbiol. 1968 Kato 1200 6Document8 paginiAppl. Microbiol. 1968 Kato 1200 6Sheerin SulthanaÎncă nu există evaluări
- Bioethics Timeline and ControversiesDocument18 paginiBioethics Timeline and ControversiesSheerin SulthanaÎncă nu există evaluări
- Human Genome ProjectDocument57 paginiHuman Genome ProjectDhaval ParekhÎncă nu există evaluări
- BCH440 (8) 32-33Document20 paginiBCH440 (8) 32-33Sheerin SulthanaÎncă nu există evaluări
- A Brief Review of Plants Having Anti Cancer PropertyDocument8 paginiA Brief Review of Plants Having Anti Cancer PropertysharammaÎncă nu există evaluări
- Chapter 26 Phylogeny and The Tree of LifeDocument41 paginiChapter 26 Phylogeny and The Tree of LifeSheerin Sulthana100% (1)
- Trade-Related Intellectual Property Rights (Trips)Document20 paginiTrade-Related Intellectual Property Rights (Trips)Sheerin SulthanaÎncă nu există evaluări
- A Business-Oriented Overview of Intellectual Property For Law StudentsDocument16 paginiA Business-Oriented Overview of Intellectual Property For Law StudentsSheerin SulthanaÎncă nu există evaluări
- Internet and NetworkingDocument42 paginiInternet and Networkingآصف رضاÎncă nu există evaluări
- Analysis On The Impact of Madrid Protocol For The Economies of Developing CountriesDocument62 paginiAnalysis On The Impact of Madrid Protocol For The Economies of Developing CountriesSheerin Sulthana0% (1)
- Radioimmunoassay: Advanced Research Techniques in Basic Medical SciencesDocument23 paginiRadioimmunoassay: Advanced Research Techniques in Basic Medical SciencesKiran FriendooÎncă nu există evaluări
- Biological Safety Levels: Endia Ford Lori Gladney Izabella OsakweDocument34 paginiBiological Safety Levels: Endia Ford Lori Gladney Izabella OsakweSheerin Sulthana100% (1)
- ATCCDocument3 paginiATCCSheerin SulthanaÎncă nu există evaluări
- Elements of Biosafety: Janet Peterson Jpeterso@accmail - Umd.edu 405-3975Document28 paginiElements of Biosafety: Janet Peterson Jpeterso@accmail - Umd.edu 405-3975Sheerin SulthanaÎncă nu există evaluări
- Dimensions of Product Quality ManagementDocument5 paginiDimensions of Product Quality ManagementSheerin SulthanaÎncă nu există evaluări
- Lecture 21 - Respiratory SystemDocument59 paginiLecture 21 - Respiratory SystemSheerin SulthanaÎncă nu există evaluări
- The Range of Quality: Quality of Design Quality of Materials Quality of ProductsDocument4 paginiThe Range of Quality: Quality of Design Quality of Materials Quality of ProductsKin SkyeÎncă nu există evaluări
- Foodborne IllnessDocument13 paginiFoodborne IllnessSheerin SulthanaÎncă nu există evaluări
- Biological DatabaseDocument19 paginiBiological DatabaseMahesh Yadav100% (8)
- Foods Poli AgeDocument23 paginiFoods Poli AgeSheerin SulthanaÎncă nu există evaluări
- Pricing ProblemsDocument2 paginiPricing ProblemsSheerin SulthanaÎncă nu există evaluări
- 06iso Allo Idio09Document18 pagini06iso Allo Idio09Sheerin SulthanaÎncă nu există evaluări
- Emerging: Brewing Is The Production ofDocument16 paginiEmerging: Brewing Is The Production ofSheerin SulthanaÎncă nu există evaluări
- 10 1371-Journal Pone 0065654 g003Document1 pagină10 1371-Journal Pone 0065654 g003Sheerin SulthanaÎncă nu există evaluări
- (WWW - Entrance Exam - Net) Animal BiotechDocument34 pagini(WWW - Entrance Exam - Net) Animal BiotechSheerin SulthanaÎncă nu există evaluări
- Antinfek 10HDocument20 paginiAntinfek 10Hmistytewest100% (1)
- 10th Standard Tamilnadu State Board Science (English Medium)Document283 pagini10th Standard Tamilnadu State Board Science (English Medium)Karthick NÎncă nu există evaluări
- 30395-Article Text-56938-1-10-20220916Document8 pagini30395-Article Text-56938-1-10-20220916Djim KARYOMÎncă nu există evaluări
- Chapter 2 Medical Terminology Verified AnswersDocument5 paginiChapter 2 Medical Terminology Verified AnswersGregg ProducerÎncă nu există evaluări
- Transport Phenomena in Nervous System PDFDocument538 paginiTransport Phenomena in Nervous System PDFUdayanidhi RÎncă nu există evaluări
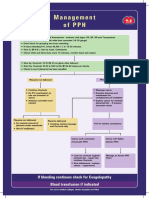
- Management of PPHDocument1 paginăManagement of PPH098 U.KARTHIK SARAVANA KANTHÎncă nu există evaluări
- 4 Factors Effecting Shelf Life and Storage Life of Fresh FishDocument26 pagini4 Factors Effecting Shelf Life and Storage Life of Fresh FishRafidah Zakaria100% (1)
- HistamineDocument16 paginiHistaminevg03ik2Încă nu există evaluări
- Cell Membrane: A Selective BarrierDocument23 paginiCell Membrane: A Selective BarrierHama JamalÎncă nu există evaluări
- Stress BreakerDocument7 paginiStress BreakerEshan Verma33% (3)
- Kingdom Protista: Diverse Eukaryotic MicroorganismsDocument79 paginiKingdom Protista: Diverse Eukaryotic MicroorganismsMeah PachecoÎncă nu există evaluări
- Adolescent Reproductive and Sexual HealthDocument42 paginiAdolescent Reproductive and Sexual HealthMuhammad Abbas WaliÎncă nu există evaluări
- 0610 IECR P3 Q1 v2 PDFDocument9 pagini0610 IECR P3 Q1 v2 PDFGAIA Educators - Home TuitionÎncă nu există evaluări
- Egg Quality Manual Web PDFDocument44 paginiEgg Quality Manual Web PDFVeterinario La EsperanzaÎncă nu există evaluări
- Classification and Characterization of Microsatellite Instability Across 18 Cancer TypesDocument13 paginiClassification and Characterization of Microsatellite Instability Across 18 Cancer TypeshadymatrixÎncă nu există evaluări
- AP Psychology Mnomonic DevicesDocument7 paginiAP Psychology Mnomonic DevicesBellony SandersÎncă nu există evaluări
- Introspect For DealersDocument13 paginiIntrospect For DealersOBERON-INTROSPECT-BIOSPECTÎncă nu există evaluări
- Molecular Cloning Technical GuideDocument40 paginiMolecular Cloning Technical GuideRÎncă nu există evaluări
- MR Spectroscopy: Technical Aspects and Clinical ApplicationsDocument5 paginiMR Spectroscopy: Technical Aspects and Clinical ApplicationsNotariana Kusuma AÎncă nu există evaluări
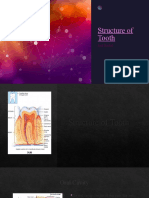
- Structure of Tooth m121Document13 paginiStructure of Tooth m121amirhossein vaeziÎncă nu există evaluări
- Biology Investigatory ProjectDocument17 paginiBiology Investigatory ProjectAnmol Dhungel100% (6)
- 4.1 - Cell Cycle Part 1Document5 pagini4.1 - Cell Cycle Part 1Deomar Joseph ParadoÎncă nu există evaluări
- Plant Transport - IGCSE Biology Notes (2020)Document6 paginiPlant Transport - IGCSE Biology Notes (2020)Zhi En LeeÎncă nu există evaluări
- Self Concept Inventory Hand OutDocument2 paginiSelf Concept Inventory Hand OutHarold LowryÎncă nu există evaluări
- Ewh Ix PDFDocument80 paginiEwh Ix PDFOR Premium FreeÎncă nu există evaluări
- Thesis Olof HerttingDocument57 paginiThesis Olof HerttingAjay IyerÎncă nu există evaluări
- Daftar PustakaDocument3 paginiDaftar PustakaAdnanÎncă nu există evaluări
- Noscapine - British PharmacopoeiaDocument3 paginiNoscapine - British PharmacopoeiaSocial Service (V)Încă nu există evaluări
- Theories of Tooth MovementDocument11 paginiTheories of Tooth MovementAhmedsy Ahmedsy AhmedsyÎncă nu există evaluări
- Short Periods of Incubation During Egg Storage - SPIDESDocument11 paginiShort Periods of Incubation During Egg Storage - SPIDESPravin SamyÎncă nu există evaluări