Documente Academic
Documente Profesional
Documente Cultură
CTO - Embriología
Încărcat de
Pablo ParicahuaDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
CTO - Embriología
Încărcat de
Pablo ParicahuaDrepturi de autor:
Formate disponibile
Manual CTO
1. edicin
Embriologa
Ecuador
A-PDF Manual Split Demo. Purchase from www.A-PDF.com to remove the watermark
No est permitida la reproduccin total o parcial de este libro, su tratamiento informtico, la transmi-
sin de ningn otro formato o por cualquier medio, ya sea electrnico, mecnico, por fotocopia, por
registro y otros medios, sin el permiso previo de los titulares del copyright.
CTO EDITORIAL, S.L. 2012
Diseo y maquetacin: CTO Editorial
C/ Francisco Silvela, 106; 28002 Madrid
Tfno.: (0034) 91 782 43 30 - Fax: (0034) 91 782 43 43
E-mail: ctoeditorial@ctomedicina.com
Pgina Web: www.grupocto.es
No est permitida la reproduccin total o parcial de este libro, su tratamiento informtico, la transmi-
sin de ningn otro formato o por cualquier medio, ya sea electrnico, mecnico, por fotocopia, por
registro y otros medios, sin el permiso previo de los titulares del copyright.
CTO EDITORIAL, S.L. 2013
Diseo y maquetacin: CTO Editorial
C/ Francisco Silvela, 106; 28002 Madrid
Tfno.: (0034) 91 782 43 30 - Fax: (0034) 91 782 43 43
E-mail: ctoeditorial@ctomedicina.com
Pgina Web: www.grupocto.es
INDICE
EMBRIOLOGA
1. Defnicin y nomenclatura .................................................................................................................................................... 1
2. Generalidades del aparato reproductor ........................................................................................................... 2
3. Gametognesis ......................................................................................................................................................................................... 2
4. Ciclo sexual femenino .................................................................................................................................................................... 4
5. Modifcaciones de los gametos previas a la fecundacin .................................................... 6
6. Fecundacin .................................................................................................................................................................................................. 7
7. Primera semana de vida ............................................................................................................................................................. 9
8. Segunda semana-implantacin .................................................................................................................................... 11
9. Tercera semana. Gastrulacin ........................................................................................................................................... 17
10. Cuarta semana ........................................................................................................................................................................................... 22
11. Desarrollo del corazn .................................................................................................................................................................. 24
12. Sisexcretor-genital ............................................................................................................................................................................... 25
13. Placenta ................................................................................................................................................................................................................ 26
14. Teratogenia ..................................................................................................................................................................................................... 26
Conceptos clave ...................................................................................................................................................................................... 27
1
EMBRIOLOGA
9
1. Denicin y nomenclatura
La embriologa es la ciencia que estudia el fenmeno de la vida antes del nacimiento, desde su formacin,
crecimiento y desarrollo hasta el parto.
El desarrollo prenatal puede subdividirse en distintos perodos, a saber (Tabla 1):
Perodo preembrionario: comprende las primeras tres semanas de vida intrauterina. Incluye los
procesos de segmentacin de la clula huevo (primera semana), implantacin en el endometrio
materno (segunda semana) y gastrulacin (tercera semana), que concluye con la formacin del
embrin trilaminar.
Perodo embrionario: abarca de la cuarta a la octava semanas de gestacin. En este perodo se for-
man los esbozos de la mayora de los rganos y sistemas del cuerpo.
Perodo fetal: desde la novena semana hasta el nacimiento, durante este perodo se terminan de
desarrollar estructural y funcionalmente los rganos. El producto de la concepcin aumenta su com-
plejidad y tambin su tamao.
Da 0 1.
a
Semana 2.
a
Semana 3.
a
Semana
4
.a
a 8.
a
Semana
9.
a
a 40.
a
Semana
Estructuras
embrionarias
Huevo o
cigoto
Blastocisto
Embrin
Bilaminar
Embrin
trilaminar
Embrin
cilndrico
Feto
Procesos
biolgicos
Fecundacin Segmentacin Implantacin Gastrulacin
Organognesis
Histognesis
Morfognesis
Desarrollo
funcional
Crecimiento
corporal
Ubicacin
Tercio distal
de la trompa
Trompa
uterina
Endometrio
(decidua)
Cuerpo
del tero
Cuerpo
del tero
Cuerpo
del tero
Perodos
Pre-
embrionario
Pre-
embrionario
Pre-
embrionario
Embrionario Fetal
Tabla 1. Desarrollo embrionario
Al referirnos al embrin o feto en el espacio utilizaremos conceptos que son similares a los empleados por
la anatoma del adulto, pero que toman palabras diferentes.
Los ejes del embrin o feto sern los mismos que los de la vida adulta; as, existe un eje mayor
o longitudinal (cfalo-caudal), uno latero-lateral (derecha-izquierda) y otro dorso-ventral (antero-
posterior).
No utilizamos en embriologa los trminos superior o inferior, ni anterior o posterior, sino que indi-
camos la ubicacin de las estructuras anatmicas como ceflicas o caudales, ventrales o dorsales,
etctera (Figura 1).
Manual CTO 1. Edicin
2
Figura 1. Anatoma embriolgica
2. Generalidades
del aparato reproductor
2.1. Aparato reproductor masculino
Los rganos sexuales pueden clasifcarse de acuerdo a su funcin en: pri-
marios y secundarios. Los rganos primarios son aquellos encargados de
la sntesis de gametos. Los rganos secundarios se encargan del trans-
porte de las mismas. De esta manera, los rganos genitales masculinos
estn formados por (Figura 2):
a) rganos sexuales primarios o gnadas: testculo.
b) rganos sexuales secundarios o sistema de conduccin:
- Va espermtica:
Conducto eferente.
Epiddimo.
Conducto deferente.
Conducto eyaculador.
Figura 2. Aparato genital masculino
- Uretra.
- rgano masculino de la cpula: pene.
- Sistema de glndulas anexas:
Prstata.
Vesculas seminales.
Glndulas bulbouretrales o de Cowpe.
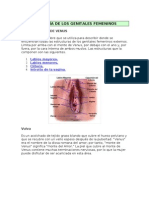
2.2. Aparato reproductor femenino
Los rganos genitales femeninos se clasifcan en (Figura 3):
a) rganos internos:
Estn representados por:
- Ovarios.
- Trompas uterinas.
- tero.
- Vagina.
b) rganos externos:
Estn representados por:
- Monte de Venus.
- Labios mayores.
- Labios menores.
- Cltoris.
Figura 3. Aparato genital femenino
3. Gametognesis
La gametognesis es el proceso por el cual se forman los gametos: es-
permatozoide u ovocito. Los gametos son clulas haploides (1n), es decir
slo tienen una copia del material gentico, de manera que cuando se
unan con el otro gameto formarn un organismo diploide (2n) (Figura 4).
El proceso de formacin de gametos se hace mediante la divisin meiti-
ca (meiosis). Las clulas que experimentan la meiosis son los espermato-
citos y ovocitos I, que se generaron a partir de la diferenciacin de esper-
matogonias y ovogonias respectivamente.
EMBRIOLOGA 9
3
Figura 4. La gametognesis
La haploida no es la nica consecuencia de la meiosis; en ella aumenta
tambin la variabilidad gentica, al intercambiarse material gentico
entre cromosomas homlogos en el crossing over y producirse la segre-
gacin de los mismos al azar (es decir que los Citos II adquieren 23 cro-
mosomas, pero stos no son todos maternos o paternos sino que consti-
tuyen una mezcla de ambos).
Las diferencias que presenta la meiosis con la mitosis se resumen en la
tabla 2.
3.1. Espermatognesis
Espermatogonias: son las clulas madre de las cuales se formarn los es-
permatozoides por un proceso denominado espermatognesis, que dura
de media 72 das. Poseen un nmero diploide de cromosomas. Pueden
dividirse (mediante mitosis) renovando la poblacin de gonias. Sin em-
bargo, algunas gonias se diferenciarn a espermatocitos I (Figura 5).
Figura 5. Mitosis y meiosis de las espermatogonias
El paso de gonias a espermatocitos I se produce por mitosis y diferen-
ciacin.
El espermatocito I, previa duplicacin del ADN, , comienza la primera di-
visin meitica (o reduccional), originando los espermatocitos II. Los
espermatocitos II experimentan la segunda divisin meitica (o ecuacio-
nal), generando las espermtides.
Las espermtides (clulas redondas) se transformarn en espermato-
zoides (clulas alargadas, con fagelo, altamente especializadas) por un
proceso de diferenciacin celular. A este proceso se lo conoce como
espermiognesis.
La espermiacion, en cambio, es la liberacin del espermatozoide a la luz
del tbulo seminfero (particularmente, de las clulas de Srtoli que com-
ponen el epitelio seminfero).
La espermatognesis, o gnesis de los espermatozoides, incluye todos
los procesos anteriores.
3.2. Espermiognesis
Conjunto de cambios que se producen en las espermtides para dife-
renciarse a espermatozoides maduros (Figura 6). Dichos cambios com-
prenden moviemientos de las organelas, disminucin del citoplasma,
condensacin de la cromatina, formacin del acrosoma y formacin del
fagelo. EL acrosoma deriva del Golgi y contiene enzimas importantes
para la fecundacin como la hialuronidasa y la acrosina.
Meiosis Mitosis
Tipo de clulas que involucra Gonias (clulas germinales) Clulas somticas
Duplicacin del ADN
nica previa a la meiosis I, es seguida de dos
divisiones del material gentico
Cada replicacin del ADN es seguida por una divisin
celular nica
Fases del ciclo celular
Fase S prolongada
Fase G2 ausente o muy breve
Fase S convencional seguida de Fase G2
Duracin
Prolongada, hasta varios aos en el sexo femenino
(ver ms adelante en ovognesis).
Breve, 1 hora aproximadamente.
Material gentico Se reduce Se mantiene constante
Crossing over
Ocurre durante el paquinema, donde se intercambia
material gentico de cada cromosoma con su
homlogo.
No ocurre, los cromosomas evolucionan de forma
independiente.
Tabla 2. Diferencias entre mitosis y meiosis
Manual CTO 1. Edicin
4
Figura 6. Espermiognesis
3.3. Ovognesis
Las clulas germinales, que en los embriones humanos aparecen en el
saco vitelino hacia la tercera semana del desarrollo, migran hacia las g-
nadas llegando a ellas al fnal de la cuarta semana. Esto sucede en uno u
otro sexo indistintamente.
Una vez instaladas en las gnadas, en caso de tratarse de un embrin feme-
nino, se transforman en ovogonias. Gran parte de dichas ovogonias siguen
dividindose por mitosis, pero algunas de ellas se diferencian a ovocitos
primarios (Figura 7). Hacia el quinto mes de vida intrauterina, el nmero
de clulas germinativas en el ovario alcanza unos 7 millones. A partir de
este momento, muchos ovogonios y muchos ovocitos primarios (aqullos
que estn ms alejados de la corteza) degeneran. Todos los ovocitos prima-
rios que han sobrevivido, ingresarn en la primera divisin meitica.
Los ovocitos I inician la meiosis intratero. La meiosis I se detiene en la
fase de diplonema o dictiotene (profase I) en el octavo mes de vida in-
trauterina. La meiosis I se reanudar en la pubertad y se detiene en meta-
fase II. Cada mes, desde la pubertad y hasta la menopausia, un grupo de
ovocitos I, reanudar su meiosis. De esto se deduce que la meiosis en el
sexo femenino puede durar de 10 aos (en el caso del primer ciclo sexual
de una nia con pubertad a esta edad) hasta ms de 40 (en los ltimos
ciclos reproductivos de una mujer pre-menopusica). Slo podr concluir
el ciclo aqul ovocito II que sea fecundado.
RECUERDA
Los ovocitos I estn detenidos en la profase I (dictiotene) hasta la pubertad.
Vale la pena mencionar las diferencias no tan sutiles existentes entre la
ovognesis y la espermatognesis. La primera comienza en el perodo
de vida prenatal y se detiene en la primera divisin meitica hasta la pu-
bertad. En ese momento se reinicia la meiosis I que se contina con la
meiosis II. sta slo terminar en caso de existir fecundacin. De ambas
meiosis se obtiene una nica gameta y 2 cuerpos polares. La esperma-
togensis, muy por el contrario, comienza en la pubertad, la meiosis I
es seguida inmediatamente por la meiosis II (el proceso completo dura
aproximadamente 24 das, 8 das para la meiosis I y 16 das para la meiosis
II) y las gametas se van renovando constantemente a lo largo del tiempo.
De un nico expermatocito I se obtienen 4 espermtides.
Figura 7. Espermatognesis y ovognesis
4. Ciclo sexual femenino
El ciclo sexual femenino involucra una serie de cambios cronolgicamen-
te ordenados y organizados por un sistema integrador. En este caso en
particular, gracias a la regulacin y la integracin que ejerce el sistema
endocrino, es posible el desarrollo de este ciclo. Al igual que cualquier
otro ciclo, no tiene ni principio ni fn, simplemente se plantea como con-
vencin su inicio con el primer da de la menstruacin y fnalizacin el
ltimo da antes de la menstruacin siguiente. Por tanto, el ciclo sexual fe-
menino es un conjunto de fenmenos que ocurren simultneamente en
el tero, trompas, ovarios y mamas (principalmente, ya que otros tejidos y
rganos tambin se adaptan) que se hallan integrados por las hormonas
del sistema endocrino y regulados por el SNC, que tiene como comienzo,
por convencin, el primer da de la menstruacin.
Se considera que un ciclo normal dura 28 das con una variacin en ms
o en menos de 7 das. Es decir, que todo ciclo que dure de 21 a 35 das
ser considerado dentro de lmites normales (Figura 8).
EMBRIOLOGA 9
5
Existe otro concepto referido a este tema, que es el de regularidad. La
regularidad habla de una variacin de un ciclo a otro de ms o menos 48
h., es decir que si una mujer tiene un ciclo de 23 das, se considera regular
si mantiene ciclos subsiguientes de entre 21 y 25 das.
El ciclo sexual femenino cumple la funcin de preparar en forma reitera-
da al aparato genital femenino para un eventual embarazo, durante lo
que se considera el perodo frtil, desde la pubertad hasta la menopausia.
Recordemos que el ciclo dura 28 das; clsicamente se lo divide en dos fa-
ses, cada una de 14 das de duracin. En el caso del ciclo sexual femenino,
el hipotlamo (parte del sistema nervioso central) produce GnRH (hormo-
na liberadora de gonadotrofnas), que estimula la hipfsis (glndula en-
dcrina). sta produce lh y fsh, que estimulan al ovario, para que produzca
estrgenos y progestgenos. Estos ltimos tienen un efecto inhibidor de la
secrecin de GnRh y de LH y FSH (actan inhibiendo en ambos niveles por
retroalimentacin negativa).
A) PRIMERA FASE (das 1 a 14)
Se denomina, a nivel uterino, fase proliferativa, porque el endometrio
est proliferando, en respuesta a la accin de los estrgenos regenerando
la capa funcional que perdi en el ciclo previo. A nivel ovrico en cambio
se conoce como fase folicular, ya que el proceso que domina el perodo
es la maduracin de los folculos ovricos.
La hormona hipofsaria que rige esta primera fase es la foliculoestimu-
lante o FSH y de las hormonas ovricas, predomina la produccin de
estrgenos con mnima secrecin de progesterona. Es por eso que esta
fase tambin puede llamarse fase estrognica. El ciclo sexual femenino
comienza, como se expresara previamente, el da de inicio de la mens-
truacin, que representa la cada de la capa funcional del endometrio en
respuesta a la isquemia (falta de oxgeno). La capa basal permanece para
proliferar en cada ciclo.
Encontrndose los niveles de estrgenos y progesterona bajos (no existe
feedback negativo), el hipotlamo libera GnRh (factor liberador de go-
nadotrofnas), seal que estimula a que la hipfsis produzca LH y FSH.
A nivel ovrico, la FSH estimula la maduracin de los folculos, en nmero
de 3 a 30 cada ciclo. Las clulas de granulosa (foliculares) producen estr-
genos que, sobre el endometrio estimularn su proliferacin. De estos 3 a
30 folculos, slo uno completar su maduracin y ser ovulado el da 14,
en respuesta a un pico de LH que se produce aproximadamente el da
13 y, aproximadamente 10-12 horas despus se produce la ovulacin
el da 14.
La LH hipofsaria tendra varias funciones:
El ovocito reanuda la meiosis I e inicia la meiosis II.
Produce la ruptura folicular y consecuente ovulacin (se elimina
el ovocito II, rodeado por una cubierta glucoproteica llamada
membrana pelcida y una capa de clulas foliculares, la corona
radiata).
Luteiniza las clulas foliculares y tecales , iniciando el cuerpo lteo la
secrecin de progesterona.
RECUERDA
La ovulacin es consecuencia del pico de LH.
El resto de los folculos que iniciaron la maduracin involucionar, convir-
tindose en folculos atrsicos.
En cuanto a la regulacin hormonal del ciclo en esta primera fase,
cabe aclarar un punto, que es crucial para entender la ovulacin. Du-
rante toda esta primera fase, los niveles hormonales que inicialmente
se encontraban bajos, fueron en aumento paulatino hasta el da 14,
teniendo los estrgenos y la progesterona un efecto de retroalimen-
tacin o feedback negativo sobre los niveles superiores (hipotlamo
e hipfsis). Sin embargo, llega un momento que se produce un pico
de estradiol que dispara el pico de LH y con ello la ovulacin, se-
guida de la luteinizacin del folculo, con secrecin de estrgenos y
progesterona (Figura 9).
RECUERDA
El ovocito es ovocito primario en profase de la 1 meiosis hasta la pubertas.
Con la ovulacin, se completa la 1 meiosis y pasa a ser ovocito secundario
hasta la fecundacin, que estimula la segunda divisin meitica.
Figura 8. El ciclo menstrual
Manual CTO 1. Edicin
6
Figura 9. Hormonas que intervienen en el ciclo sexual femenino
B) SEGUNDA FASE (das 15 a 28)
A nivel uterino se conoce como fase secretoria ya que el endometrio pre-
senta un estado de activa secrecin de sus glndulas. A nivel ovrico es
la fase ltea, ya que es el cuerpo lteo la estructura que permanece en el
rgano despus de la ovulacin. La hormona hipofsaria que predomina
es la LH, que estimular a que el cuerpo lteo produzca progesterona.
Esta fase es entonces conocida tambin como progestacional.
El cuerpo lteo produce estrgenos y progesterona, siendo esta ltima quien
convierte el endometrio, previamente proliferado por los estrgenos, en se-
cretor. La secrecin endometrial es importante ya que en caso de existir fecun-
dacin, la clula huevo deber nutrirse de las secreciones uterinas (y tubarias)
hasta que se produzca la adecuada implantacin y se forme la placenta. El
cuerpo lteo inicia su involucin unos 10 das posteriores a la ovulacin, co-
menzando a disminuir los niveles de estrgenos y progesterona en sangre.
Algo a destacar en este punto es que el cuerpo lteo tiene una vida me-
dia de 14 das, lo que hace que esta fase del ciclo se mantenga constante.
Es por eso que para calcular la fecha de ovulacin de una mujer, debe
restarse a la cantidad de das que dure su ciclo, 14. En un ciclo de 28 das
la ovulacin se produce el da 14 (justo en la mitad del mismo) pero en un
ciclo de 34 das, se producir el da 20 (34 - 14 = 20).
RECUERDA
EL cuerpo lteo tiene una vida media de 14 das, por lo que la segunda fase
del ciclo es constante.
En caso de ocurrir la fecundacin, la evolucin del cuerpo lteo sera dis-
tinta. Considerando que la fecundacin ocurre cercana a la ovulacin,
aproximadamente el da 14, el huevo o cigoto se traslada por la trompa
hacia la cavidad uterina, llegando a sta unos 4 das ms tarde. Aproxima-
damente el da 7 de vida (21 del ciclo sexual femenino), se inicia la implan-
tacin del huevo en el endometrio materno. Es en este momento en que
se inicia la secrecin (por parte de las clulas que intervienen en la implan-
tacin) de una nueva hormona, la GCH o HCG (gonadotrofna corinica
humana), similar a la LH en estructura y funcin. Ser ella la responsable
de estimular al cuerpo lteo para que contine secretando progesterona,
para mantener el endometrio en fase secretora. En este caso, el cuerpo
lteo permanece viable hasta la semana 12 de gestacin, momento en
que la placenta produce progesterona en cantidades adecuadas.
5. Modicaciones de los gametos
previas a la fecundacin
5.1. Caractersticas
del ovocito despus de la ovulacin (Figura 10)
Como se expusiera previamente, el ovocito II (detenido en metafase de la
meiosis II), es expulsado del ovario rodeado por una capa de clulas foli-
culares, la corona radiata. Entre la corona radiata y la membrana plasmtica
del ovocito se encuentra la membrana pelcida, compuesta por glucopro-
tenas. A nivel de la membrana pelcida se ubican molculas importantes
que permitirn la interaccin con el espermatozoide. stas son ZP1, ZP2 y
ZP3 (ZP por zona pelcida).
El ovocito posee asimismo numerosas microvellosidades, que atraviesan la
membrana pelcida y contactan con las clulas foliculares de la corona. En el
citoplasma reconocemos un sector altamente diferenciado, la periferia ovu-
lar, que posee una densa trama de microflamentos y flamentos interme-
dios, junto con vesculas corticales (de ah que a esta zona perifrica se le d
tambin el nombre de corteza). stas poseen un contenido enzimtico y se
encuentran involucradas en el bloqueo de la polispermia. El resto de las or-
ganelas se ubican ms profundamente en el citoplasma, cercanas al ncleo.
Figura 10. Ovocito II post-ovulacin
5.2. Modicaciones que se producen en los
espermatozoides previos a la fecundacin
A) MADURACIN
Es el proceso que engloba todas las modifcaciones preparatorias que sufre
el espermatozoide en el tracto genital masculino (TGM), relacionadas con la
adquisicin de capacidad fecundante. El rgano clave en este proceso es el
epiddimo, ya que a nivel de su cabeza y cuerpo se producen los cambios
que son ms de carcter funcional y bioqumico que estructurales.
Los objetivos de esta etapa son:
1. Que las molculas de la superfcie del espermatozoide que actuarn
como receptores en la fecundacin, se encuentren enmascaradas.
EMBRIOLOGA 9
7
2. Disminuir la actividad del espermatozoide, para reservar la energa
del mismo.
3. Estabilizar la membrana plasmtica.
Para que estos espermatozoides ya maduros (que se encuentran almace-
nados en la cola del epiddimo) adquieran capacidad fecundante necesi-
tan un paso ms: la capacitacin.
B) CAPACITACIN
Es el conjunto de modifcaciones bioqumicas previas a la fecundacin
que ocurren en el tracto genital femenino (TGF). Los fuidos del TGF seran
altamente capacitantes, principalmente en el perodo periovulatorio. No
se considera crucial la actividad de un rgano femenino en particular sino
que los cambios ocurriran a medida que el espermatozoide asciende des-
de la vagina hasta su encuentro con el ovocito II en la trompa de Falopio.
Incluye procesos que pueden en parte considerarse opuestos a los ocu-
rridos en la maduracin.
Los cambios en los lpidos de membrana seran responsables de un au-
mento de su capacidad fusgena, que como se ver ms adelante es im-
portante para uno de los pasos de la fecundacin, la reaccin acrosmica,
y, por supuesto, para la ulterior fusin de la membrana del espermatozoi-
de con la del ovocito II.
RECUERDA
La maduracin ocurre en el tracto genital masculino, mientras que la capa-
citacin ocurre en el tracto genital femenino.
6. Fecundacin
La fecundacin es un hecho sumamente complejo y excepcional de la
biologa que permite la fusin de dos clulas de distintos individuos (ga-
metos), dando como resultado un nuevo organismo unicelular diploide, la
clula huevo, impulso inicial para el desarrollo de un nuevo ser (Figura 11).
Figura 11. Fases de la fecundacin
6.1. Transporte de gametos
Durante la ovulacin, el ovocito fecundante es transferido a un nuevo
medio, las trompas de Falopio, y desde all se dirige lentamente hacia la
cavidad uterina.
El transporte del espermatozoide por las vas de conduccin masculinas
es netamente pasivo, siendo la fuerza muscular de las paredes del TGM la
que impulsa los espermatozoides. En el TGF, stos son depositados en el
fondo de saco vaginal posterior. Atraviesan el cuello uterino y comienzan
a ascender por la cavidad uterina en direccin a las trompas. Aqu el as-
censo puede considerarse en parte pasivo, por la actividad muscular pe-
ristltica y al movimiento coordinado de las cilias, presentes en el epitelio
de revestimiento del tracto genital femenino, y tambin activo, merced
a los movimientos del fagelo (estos movimientos son ms importantes
para atravesar el moco cervical y llegar al tero) (Figura 12).
Figura 12. transporte de gametos en el tracto femenino
6.2. Capacitacion del espermatozoide
Como consecuencia de los cambios bioqumicos que sufre el espermato-
zoide, la actividad metablica del espermatozoide abandona el letargo y
aumenta signifcativamente: el consumo de O
2
es considerablemente ma-
yor, el fagelo se activa y hay importantes cambios en la composicin de
fosfolpidos y partculas intramembranosas (esto vuelve a la membrana
sumamente inestable, aumentando su capacidad fusgena) (Figura 12).
6.3. Encuentro de los gametos
El encuentro de los gametos se produce en el tercio distal de la trompa de
Falopio, a nivel de la unin istmo - ampular de la misma. El ovocito tarda
unos pocos minutos en llegar al punto de encuentro despus de la ovula-
cin. De los 300 a 400 millones de espermatozoides depositados en la vagina
durante la eyaculacin, slo unos pocos sobrevivirn al medio relativamente
hostil en que se encuentran y llegarn sanos y salvos al lugar de encuentro.
RECUERDA
La fecundacin se produce en el tercio distal de la trompa de Falopio.
Manual CTO 1. Edicin
8
6.4. Penetracin de la corona radiata
Las clulas foliculares de la corona se encuentran unidas entre s por ci-
do hialurnico. El espermatozoide toma contacto con la corona radiata y
la penetra, gracias a la fuerza impulsora generada por los movimientos de
hiperactivacin. Va labrando un tnel entre las clulas foliculares, merced
a la accin enzimtica de la hialuronidasa que reviste la superfcie exter-
na del espermatozoide (idntica a la hialuronidasa que se encuentra en
la vescula acrosmica). Despus de penetrar la corona, la superfcie del
espermatozoide toma contacto con la membrana pelcida.
6.5. Reconocimiento y reaccin acrosmica
La protena ZP3 de la membrana pelcida interacta con una galactosil-
transferasa de la membrana post-acrosmica del espermatozoide, des-
encadenando una cascada de eventos intracelulares en el espermatozoide
que llevarn a la reaccin acrosmica (Figura 13). Este reconocimiento es
especfco de especie, es decir, que diferentes molculas se reconocen en las
diferentes especies, previniendo de esta forma la fusin de gametas de espe-
cies distintas y consecuente generacin de organismos hbridos (Figura 14).
RECUERDA
En el reconocimiento de las gametas intervienen ZP3 del ovocito y una ga-
lactosiltransferasa del espermatozoide.
Al inicio de la reaccin acrosmica se observa un fenmeno bastante
raro en la biologa, la fusin de membranas. La membrana plasmtica
del espermatozoide se fusiona, a intervalos ms o menos regulares, con
la membrana externa del acrosoma, originando pequeas aberturas
o poros que permiten la liberacin de las enzimas. Las zonas donde las
membranas no se fusionan dan una imagen de vesculas al microscopio
electrnico, de ah la denominacin de proceso de vesiculizacin a esta
primera etapa de la reaccin acrosmica. Finalmente, las vesculas se des-
prenden y dejan expuesta la membrana interna del acrosoma con sus
partculas fjadas en ella.
Figura 13. La reaccin acrosmica.
Figura 14. Reconocimiento de gametos
EMBRIOLOGA 9
9
La reaccin acrosmica tiene como consecuencias:
La denudacin o desaparicin de la corona radiata por la accin de
la hialuronidasa liberada del acrosoma.
La penetracin de la membrana pelcida por accin de la acrosina:
la membrana pelcida debe permanecer hasta el sexto da de gesta-
cin, es decir, hasta momentos antes del inicio de la implantacin. En
esta fase se penetra pero no se destruye.
La fusin de las membranas plasmticas de ambas gametas.
RECUERDA
La acrosina es responsable de la penetracin de la membrana pelcida. La
acrosina se libera como un pro-enzima que al contactar con la ZP2 de la
zona pelcida se activa.
6.6. Activacion del ovocito
Se defne de este modo a la serie de cambios biomoleculares que se
desarrollan en el ovocito al entrar en contacto con el espermatozoide.
En esta etapa de la fecundacin se pone en marcha toda una comple-
ja maquinaria molecular que hasta entonces se mantena en estado de
latencia, y que permite el inicio del programa del desarrollo temprano
del futuro ser. El encuentro y fusin de los gametos desencadena una
serie de seales moleculares dentro del ovocito que, en ltima instancia,
producirn cambios citoplasmticos y nucleares. Puede afrmarse que el
espermatozoide enciende el interruptor que permite la activacin del
ovocito.
Clsicamente los eventos relacionados con la activacin se dividen en
tempranos o tardos. Los primeros son los ocurridos durante los pri-
meros cinco minutos posteriores al contacto espermatozoide-ovocito
(E-O).
1. Eventos tempranos de la activacin:
a) Bloqueo rpido de la polispermia: (entrada masiva de Na+)
en los primeros 3 segundos tras el contacto E-O hay una rpi-
da apertura de canales de sodio en la membrana del ovocito yse
produce una despolarizacin de la membrana. Este fenmeno
elctrico impide el ingreso de nuevos espermatozoides, consti-
tuyendo el denominado bloqueo rpido de la polispermia.
RECUERDA
El bloqueo rpido de la poliespermia depende del ingreso de Na+, mien-
tras que el bloqueo lento depende de la entrada de Ca++ y la consecuente
reaccin cortical.
b) Bloqueo lento de la polispermia: (reaccin cortical) a los 20
segundos de ocurrido el contacto E-O, se activa la liberacin de
Ca++ al citoplasma desde los depsitos intracelulares (retculo en-
doplsmico, mitocondrias, etc.). El in calcio es el responsable del
proceso de exocitosis gracias al cual son liberadas hacia el espacio
extracelular (tambin llamado perivitelino) las enzimas contenidas
en las vesculas corticales del ovocito. stas modifcan las caracte-
rsticas fsico-qumicas del espacio perivitelino, impidiendo en for-
ma defnitiva el ingreso de nuevos espermatozoides.
Como consecuencia de la reaccin cortical la membrana pel-
cida termina desprendindose de la ovocitaria y aparece entre
ambas una pequea cavidad, el espacio de fertilizacin.
2. Eventos tardos de la activacin:
Aparentemente el aumento del calcio citoplsmico sera el desenca-
denante de numerosos segundos mensajeros intracelulares que con-
ducirn a los eventos propios de la activacin tarda.
La activacin tarda incluye:
- El aumento de la sntesis proteica
- La culminacin de la meiosis II del ovocito, de la que se obtiene
un segundo cuerpo polar y el vulo que ya es en realidad la clula
huevo, por compartir el citoplasma con el material gentico del
espermatozoide.
- La formacin de los proncleos femenino y masculino: el mate-
rial gentico de cada progenitor se coloca en el centro de la clu-
la huevo, se desenrolla la cromatina y se duplica el ADN (fase S
de la primera mitosis de la clula huevo).
- La singamia y anfmixis. Se llama singamia a la prdida de las en-
volturas nucleares de los proncleos (cariotecas) y anfmixis a la
ubicacin de los cromosomas, apareados, en el plano ecuatorial,
conformando la metafase de la primera mitosis (primera divisin
de segmentacin de la clula huevo).
La anfmixis es considerada el ltimo paso de la fecundacin y el pri-
mero de la segmentacin de la clula huevo. La segmentacin ocurrir
durante toda la primera semana de gestacin, a medida que el cigoto
se traslada por la trompa de Falopio hacia la cavidad uterina.
6.7. Consecuencias de la fecundacin
1. RESTABLECIMIENTO DEL NMERO DIPLOIDE DE CROMOSOMAS.
Con la fecundacin, se obtiene el nmero de 46 cromosomas, 23 cromo-
somas provenientes del espermatozoide, y 23 cromosomas provenientes
del ovocito II.
2. DETERMINACIN DEL SEXO CROMOSMICO.
Al concluir la fecundacin, se produce la determinacin del sexo a ex-
pensas del cromosoma sexual materno, que siempre es X, y el cromo-
soma sexual paterno, que puede ser X o Y. Queda claro, por ende, que la
determinacin del sexo depende del cromosoma sexual presente en el
espermatozoide.
RECUERDA
El sexo depende del cromosoma sexual paterno (X o Y).
3. FORMACIN DE UN NUEVO SER E INICIO DE LA SEGMENTACIN.
7. Primera semana de vida
La fecundacin, en la cual se unieron un ovocito II y un espermatozoide,
culmina con la anfmixis, que consiste en la unin de los proncleos fe-
menino y masculino en la metafase correspondiente a la 1.
a
divisin de
segmentacin.
Manual CTO 1. Edicin
10
La clula huevo o cigoto (as se llama al embrin unicelular), posee su
material cromosmico ya duplicado, con informacin gentica tanto ma-
terna como paterna, y se encuentra en la trompa de Falopio (tercio distal),
todava rodeada por la membrana pelcida (PAS+, de origen glucopro-
teico), de la cual se desprender hacia el sexto da de vida embrionaria
(Figura 15). Una vez que el ovocito fecundado termina la meiosis, los cro-
mosomas maternos y paternos se unen para formar el cigoto, que contie-
ne un nico ncleo diploide. Se considera que el desarrollo embrionario
comienza a partir de este momento.
Figura 15. Clula huevo o cigoto
7.1. Segmentacin
El embrin que se acaba de formar comienza a realizar una serie de divi-
siones llamadas segmentaciones en su camino desde el tercio distal de
la trompa hasta el tero. Se trata de divisiones mitticas formndose dos
clulas, posteriormente cuatro, luego ocho y as sucesivamente. En estas
divisiones, el citoplasma de las clulas que se van formando no crecen,
por lo que el tamao del embrin no cambia.
RECUERDA
La segmentacin hace que el cigoto se divida sin aumentar de tamao.
Cuando el embrin tiene entre 8 y 16 clulas comienzan a diferenciarse
dos agrupaciones de clulas: una masa celular interna (embrioblasto, que
dar lugar al embrin y al amnios) y una externa (trofoblasto que dar
lugar a la placenta y las membranas). Este proceso por el cual una clula
decide si ser del macizo interno o externo es una determinacin, en
este caso la primera determinacin.
El embrin contina dividindose y cuando tiene 30 clulas se denomina
mrula, esto coincide con el cuarto da de gestacin.
Hacia el quinto-sexto da de desarrollo embrionario, el embrin tiene
unas 100 clulas y se denomina blastocisto (Figura 16). Se va formando
una cavidad por la incorporacin de iones y consecuentemente agua,
esta cavidad se denomina cavidad del blastocisto o blastocele.
El MCI (macizo celular interno o embrioblasto) es desplazado a uno de los
polos del blastocisto, que de ahora en adelante llamaremos polo embrio-
nario (y al polo opuesto, anembrionario).
Hacia el quinto da de gestacin, el blastocisto pierde la membrana pe-
lcida y procede a implantarse. Un da ms tarde se fja al epitelio endo-
metrial por su polo embrionario y rpidamente el trofoblasto comienza
a proliferar, generando dos tejidos diferentes: el sincitiotrofoblasto, en
contacto con el epitelio endometrial, y el citotrofoblasto, rodeando al
embrin y separndolo de la cavidad uterina.
Figura 16. Blastocisto
RECUERDA
El embrin se implanta en forma de blastocisto el 5-6 da de gestacin.
Las clulas del MCI que estn en contacto con el blastocele forman el hipo-
blasto o endodermo primitivo. As queda constituido el embrin unilaminar.
Durante su viaje hacia el tero, la nutricin del blastocisto estuvo a car-
go de las secreciones de la trompa y de las glndulas uterinas (que se
encuentran en fase secretoria). Los nutrientes ingresaron a ste por difu-
sin. Hablamos en esta primera semana de nutricin mucotrofa.
La membrana pelcida ha estado en ntima relacin con el embrin du-
rante esta primera semana de gestacin y se ha eliminado 24 horas antes
de la implantacin. Resumiendo, sus funciones han sido:
Reconocimiento especie especfico con el gameto masculino duran-
te la fecundacin (interviene la mlecula ZP3).
Bloqueo de la polispermia (despus de la reaccin cortical, la ZP3 se-
ra enzimticamente modificada, no pudiendo producirse un nuevo
reconocimiento con otros espermatozoides).
Filtro nutritivo: los nutrientes que deben difundir desde las secrecio-
nes maternas son bioqumicamente filtrados por dicha membrana.
Barrera inmunolgica: la membrana pelcida carecera de molculas
antignicas capaces de provocar un rechazo materno del embrin.
Soporte mecnico de las blastmeras: impide que las mismas se di-
socien.
Impide la implantacin prematura del embrin. La membrana pelci-
da se pierde justo antes de producirse la implantacin.
7.2. Patologa de la primera semana
Abortos: el embrin podra ser abortado espontneamente sin sa-
ber la madre que estuvo embarazada (el embrin tiene 7 das cuando
EMBRIOLOGA 9
11
la madre se encuentra en el da 21 de su ciclo, lo cual quiere decir
que es muy posible que ella no sospeche su embarazo). Un aborto
temprano puede ser causado por:
- Agentes txicos ingeridos por la madre, que ingresaran al em-
brin a travs de su nutricin (secreciones uterinas).
- Fecundacin ocurrida entre gametas viejas o portadoras de algu-
na malformacin gentica importante.
Formacin de mosaicos: si ocurre no disyuncin de un par de cromo-
somas durante la segmentacin, se obtendr un embrin con dos o
ms lneas celulares, con nmeros cromosmicos diferentes (Figura 17).
Lo normal sera que de una clula huevo de 46 cromosomas se origi-
naran siempre dos hijas de 46 cromosomas cada una. Si se forma un
mosaico, se obtienen dos hijas de 45 y 47 cromosomas cada una.
El mosaicismo es una de las causas de sndrome de Down, en la que
un embrin con 46 cromosomas (normal) realiza una no disyuncin
mittica en la segmentacin, de la que se obtienen clulas con 47
cromosomas y trisoma 21 (y por supuesto otras con 45 cromosomas
y monosoma 21). Otras blastmeras hacen sus mitosis normalmente,
obtenindose entonces un embrin compuesto por clulas normales
y otras con trisoma.
8. Segunda semana-implantacin
La implantacin es el proceso mediante el cual, durante la 2. semana de
gestacin, el embrin se introduce en el endometrio materno (Figura 18).
Con fnes didcticos describiremos inicialmente los cambios ocurridos
en el trofoblasto, referidos a la implantacin propiamente dicha y luego
los cambios en el embrioblasto, aunque debe comprenderse que ambos
procesos son simultneos.
Lugar: la implantacin normal se produce generalmente sobre la pa-
red posterior del tero materno, en su tercio superior, aunque se con-
sidera normal la implantacin que se lleve a cabo sobre cualquiera
de las paredes uterinas. Esto es lo que se conoce como implantacin
eutpica (eu= verdadero), es decir, en el lugar correcto.
Orientacin: el embrin se orienta con el polo embrionario (PE) ha-
cia el endometrio.
Tipo: la implantacin en el humano es de tipo intersticial, ya que el
embrin se introduce completamente en el espesor del endometrio
materno (Figuras 19 y 20).
Figura 18. Blastocisto antes de implantarse
RECUERDA
La implantacin es eutpica si se realiza en las paredes uterinas y ectpica
si lo hace en un sitio diferente.
Figura 17. Formacin de mosaicos
Manual CTO 1. Edicin
12
Figura 19. Blastocisto implantndose
Figura 20. Blastocisto implantado
8.1. Evolucin del trofoblasto
Podemos dividir el proceso de implantacin en fases o etapas, stas son:
Humoral: es previa al contacto fsico. En ella el embrin y el endome-
trio interactan a distancia. La secrecin de enzimas por el trofoblasto
y por el endometrio permiten la disolucin de la membrana pelcida y
el blastocisto se orienta con el polo embrionario hacia el endometrio.
De contacto propiamente dicho: consiste en la aposicin blasto-
cisto-endometrio, y es posterior a la degradacin de la membrana
pelcida.
Invasin del endometrio: comienza la penetracin del endometrio por
parte del trofoblasto, generndose dos nuevos tejidos a partir de ste:
- El sincitiotrofoblasto, una masa citoplsmica multinucleada en
contacto con el endometrio materno.
- El citotrofoblasto, formado por clulas cbicas mononucleadas,
queda rodeando al blastocisto.
RECUERDA
Del trofoblasto se generan el citotrofoblasto y el sincitiotrofoblasto.
La actividad proliferativa queda a cargo del citotrofoblasto (slo en l se
han registrado mitosis). As, parte de sus clulas se van diferenciando en
sincitiotrofoblasto.
La penetracin del blastocisto en el endometrio queda a cargo del sinci-
tiotrofoblasto. El mecanismo de penetracin no implica la destruccin
del tejido, sino que las clulas del sincitiotrofoblasto se intercalan con las
del endometrio. Este proceso de invasin particular se conoce como in-
trusin.
RECUERDA
El sincitiotrofoblasto es responsable de la invasion del endometrio.
Una vez que el embrin se ha introducido completamente en el endo-
metrio (das 11-12), se forma un cogulo de fbrina sobre el lugar donde
comenz la implantacin, que repara la superfcie uterina, quedando el
embrin atrapado en tejido materno (implantacin intersticial). La nutri-
cin durante la segunda semana de vida puede considerarse inicialmen-
te histotrofa, ya que el embrin se nutre de sustancias obtenidas del teji-
do (histos) endometrial. En el sincitiotrofoblasto aparecen luego espacios
que se llenan de plasma materno y secreciones de glndulas endome-
triales erosionadas: son las lagunas sincitiales. La circulacin por las la-
gunas se conoce como circulacin uteroplacentaria primitiva (Figura 21).
Hablamos, a fnes de esta segunda semana, de nutricin hemotrofa, si
bien se sabe que las lagunas no contienen sangre entera (con glbulos
rojos) sino un trasudado del plasma que permite la llegada al embrin
de elementos nutritivos. Los nutrientes contenidos en la sangre se trans-
portan por difusin hacia el embrin (la sangre materna recin invade la
placenta en la semana 12).
Figura 21. Etapa lacunar del desarrollo embrionario
RECUERDA
La nutricin en la segunda semana es histotrofa y hemotrofa.
Por ltimo, a partir del da 12 aproximadamente, comienzan a brotar del
citotrofoblasto, cordones celulares que crecen hacia el sincitio, constitu-
yendo las vellosidades primarias (Figura 22).
EMBRIOLOGA 9
13
RECUERDA
Las vellosidades primarias se encuentran formadas por un eje de citotrofo-
blasto rodeado de sincitiotrofoblasto.
Figura 22. Vellosidades primarias
8.2. Evolucin del embrioblasto
o MCI (macizo celular interno)
8.2.1. Formacin de la cavidad amnitica-epiblasto
Nuestro embrin de 7 das, o blastocisto, se encuentra formado por el tro-
foblasto, de ubicacin perifrica, y el macizo celular interno o embrioblasto,
adyacente al polo embrionario (Figura 23). Su cavidad es el blastocele, y en
contacto con ella aparece hacia fnes de la primera semana el hipoblasto.
As queda constitudo el embrin unilaminar.
Figura 23. Blastocisto
Alrededor del da 8, comienza a aparecer lquido entre las clulas del MCI,
formndose una nueva cavidad, la cavidad amnitica (Figura 24). El MCI
queda entonces fraccionado en dos sectores: por un lado, las clulas que
quedan entre la cavidad y el citotrofoblasto, toman el nombre de amnio-
blastos y forman una delgada membrana llamada amnios; en cambio, las
clulas que quedan apoyadas sobre el hipoblasto o endodermo primitivo
constituyen el epiblasto (o ectoblasto). Del epiblasto van a derivar la tota-
lidad de los tejidos intraembrionarios y tambin algunos extraembrionarios.
Figura 24. Formacin de la cavidad amnitica
RECUERDA
El epiblasto, hipoblasto y el amnios derivan del embrioblasto.
A fnes de la primera semana y principios de la segunda se produce la se-
gunda determinacin. Consiste en la determinacin del embrioblasto
en dos sentidos evolutivos diferentes:
Hipoblasto.
Epiblasto.
El estmulo determinante sera el medio ambiente local. As, las clulas
del MCI en contacto con el blastocele, se determinan a hipoblasto, mien-
tras que aqullas que no estn en contacto con ste, toman otra va evo-
lutiva, y se determinan a epiblasto.
8.2.2. Formacin del mesodermo extraembrionario y
celoma extraembrionario
Un grupo de clulas del hipoblasto migra rodeando al antiguo blastoce-
le. Esta cavidad es ahora el saco vitelino primitivo o cavidad exocelmi-
ca, y la membrana que lo recubre, membrana de Heuser o membrana
exocelmica.
Hacia el da 11, el embrin ha penetrado completamente en el endome-
trio materno. Un grupo de clulas de la parte ms caudal del epiblasto
Manual CTO 1. Edicin
14
migra para ubicarse por dentro del citotrofoblasto y por fuera de la cavi-
dad amnitica y saco vitelino primitivo, constituyendo un nuevo tejido, el
mesodermo extraembrionario.
RECUERDA
El mesodermo extraembrionario deriva del epiblasto.
Resumiendo lo visto hasta ahora, tenemos dos cavidades, la amnitica
y el saco vitelino primitivo, separadas por el disco embrionario bilami-
nar (formado por el epiblasto y el hipoblasto). La cavidad amnitica est
tapizada por el amnios y el saco vitelino primitivo, por la membrana de
Heuser.
En el mesodermo extraembrionario comienzan a aparecer cavidades,
que se van uniendo, coalescen, y forman el celoma extraembrionario o
cavidad corinica (un celoma es simplemente un espacio).
Decimos que el mesodermo extraembrionario se delamina (queda
constitudo por dos lminas). La lmina que rodea al saco vitelino se
llama hoja visceral o esplcnica del mesodermo extraembrionario. La
lmina que rodea a la cavidad amnitica y que tapiza al citotrofoblas-
to por dentro se llama hoja parietal o somtica del mesodermo extra-
embrionario (Tabla 3).
La nica parte del mesodermo extraembrionario que no se delamina
constituye el pedculo de fjacin o tallo de conexin, que mantiene al
embrin colgando dentro del celoma extraembrionario. La parte de la
hoja parietal que recubre la cavidad amnitica se contina con la que recu-
bre al citotrofoblasto pasando por el pedculo (ver grfco en esta pgina).
Hoja parietal Citotrofoblasto
Mesodermo ee Cav. amnitica
Hoja visceral Saco vitelino
Tabla 3. Mesodermo extraembrionario
Una nueva oleada de clulas del hipoblasto migra renovando el reves-
timiento del saco vitelino, desplazando la membrana de Heuser. El saco
vitelino, con su nuevo revestimiento endodrmico (recordemos que el
hipoblasto es un endodermo primitivo), es ms pequeo, y toma el
nombre de saco vitelino secundario o defnitivo. El remanente del saco
primitivo se reconoce como un quiste en el polo anembrionario, el quiste
exocelmico, que normalmente involuciona.
La pared del saco vitelino defnitivo se encuentra entonces formada
por dos tejidos (Figura 25):
Endodermo extraembrionario (por dentro).
Mesodermo extraembrionario hoja visceral (por fuera).
El mesodermo extraembrioario hoja parietal, el citotrofoblasto y el sinci-
tiotrofoblasto forman una membrana llamada corion, que se encuentra
por fuera del celoma extraembrionario o cavidad corinic.
Del endodermo del saco vitelino surge la alantoides, una evaginacin
hueca que se introduce en el espesor del pedculo o tallo de fjacin. Si
bien se desconoce su funcin en los humanos, se cree que induce la for-
macin de los vasos alantoideos (que posteriormente darn origen a los
vasos umbilicales).
Figura 25. Formacin del saco vitelino primario
Mientras, en el MCI aparece la cavidad amnitica, y quedan forma-
dos el amnios y el epiblasto. Se forma entonces el embrin bilami-
nar (epiblasto-hipoblasto). Se establece en este momento el eje
dorso-ventral del embrin, es decir, de ahora en ms sabemos que
el epiblasto dar origen a la parte dorsal del embrin, mientras que el
hipoblasto, si bien no da derivados intraembrionarios, es el lado ven-
tral. Los ejes cfalo-caudal y derecha-izquierda se establecern en la
tercera semana.
RECUERDA
El eje dorso-ventral se establece en la segunda semana de vida y el eje cfa-
lo-caudal se establece en la tercera semana de gestacin.
El blastocele es ahora el saco vitelino primitivo, rodeado por un epitelio
de origen hipoblstico, la membrana de Heuser o exocelmica.
Del epiblasto migran unas clulas, que se interponen entre el disco em-
brionario y el citotrofoblasto: estas clulas forman el mesodermo extra-
embrionario. ste luego se delamina, formando las hojas parietal (so-
mtica) y visceral (esplcnica).
Otras clulas endodrmicas (hipoblsticas) luego migran y tapizan este
saco vitelino, que desde ese momento se llamar defnitivo. Su pared
se encuentra formada por endodermo y mesodermo extraembrionario
hoja visceral.
RECUERDA
Las clulas geminales primordiales se originan en el interior del ectodermo
primitivo del embrin y a continuacin emigran hacia el saco vitelino.
La cavidad amnitica queda revestida por el amnios y, por fuera de ste,
la hoja parietal del mesodermo extraembrionario.
EMBRIOLOGA 9
15
Como conclusin podemos decir que nos queda un embrin bilami-
nar, rodeado por dos cavidades, la amnitica y el saco vitelino, conec-
tado por el pedculo de fjacin al corion, una membrana que rodea
al embrin en su totalidad, y que est compuesta por tres tejidos:
mesodermo extraembrionario hoja parietal, cito y sincitiotrofoblasto.
Slo existen las vellosidades primarias, compuestas por cito y sincitio-
trofoblasto.
RECUERDA
Llamamos a la SEGUNDA SEMANA la semana de LOS DOS:
Del trofoblasto se generan dos tejidos: cito-sincitio.
Dos cavidades: amnitica y saco vitelino.
Embrin bilaminar: epiblasto-hipoblasto.
El mesodermo extraembrionario se delamina en dos hojas: parietal y
visceral.
8.3. Reaccin decidual
Forman parte de la reaccin decidual los cambios que experimenta el
endometrio debidos a la presencia del embrin en la cavidad uterina. El
endometrio fomenta la invasin llevada a cabo por el sincitiotrofoblasto,
facilitando as la implantacin. Por eso decimos que la implantacin es
el resultado de la interaccin recproca entre el embrin y el endo-
metrio.
Podemos hablar de tres fases, aunque slo las dos ltimas forman parte
de la reaccin decidual:
Reaccin pseudodecidual o fase progestacional: corresponde a
la segunda parte del ciclo sexual femenino y se presenta en cada
ciclo, sin importar si hubo o no fecundacin (por eso es pseudo-
decidual). Consiste en la presencia de un endometrio secretor,
edematoso, apto para la implantacin. La hormona progesterona,
responsable de tornar el endometrio en secretor, tiene su pico plas-
mtico el da 21 (da 7 de vida embrionaria, es decir, coincidente con
el momento en que ocurre la implantacin).
Reaccin decidual primaria: corresponde a la fase preimplantato-
ria del blastocisto, cuando ste no ha tomado contacto con la pa-
red uterina. Las clulas del estroma endometrial se decidualizan,
se tornan polidricas, plidas por el acmulo intracelular de gluc-
geno y lpidos, aumentan de tamao. Puede observarse adems la
presencia de glndulas tortuosas, abundantes en secreciones, y un
estroma edematoso. Estos cambios ocurren en el endometrio adya-
cente al blastocisto y seran desencadenados por sustancias libera-
das por ste.
Reaccin decidual secundaria: ocurre a partir del contacto fsico
blastocisto-endometrio. En esta fase aumenta el nmero de clu-
las deciduales y aparece un infiltrado inflamatorio linfocitario en el
estroma endometrial. Los cambios, inicialmente locales, difunden
a todo el endometrio, que de ahora en adelante se llamar deci-
dua.
8.4. Test de embarazo-hormonas
El sincitiotrofoblasto, desde su formacin, comienza a secretar HCG (go-
nadotrofna corinica humana, hormona de tipo glucoproteico), similar
a la LH hipofsaria. La funcin de la HCG ser la de mantener el cuerpo
lteo para que siga secretando progesterona. La progesterona mantiene
al endometrio en fase secretora, imprescindible para la continuacin del
embarazo. La extirpacin quirrgica del ovario (con el cuerpo lteo ges-
tacional) antes de la semana 12 de embarazo, provoca el aborto, ya que
disminuye la secrecin endometrial por la reduccin de la progesterona
plasmtica.
RECUERDA
La funcin de la HCG es estimular al cuerpo lteo para que secrete proges-
terona.
La HCG posee dos subunidades, una alfa, similar en estructura a la LH,
que es la que determina la funcin hormonal, y una subunidad beta, di-
ferente, exclusiva de la HCG. Es decir, que de existir embarazo, el dosaje
de subunidad beta ser positivo, mientras que dosar HCG total podra dar
resultados falsos positivos (porque puede por error confundirse con la LH
ya que comparten la sub-alfa).
La deteccin de beta-HCG: el test de embarazo tradicional detecta mo-
lculas de HGD en la orina, y es el mtodo ms utilizado, permitiendo
diagnosticar la gestacin a partir de la 4-5 semana de amenorrea. Tam-
bin es posile determinar la HCG en sangre a partir de la implantacin (3
semana de amenorrea) siendo el mtodo diagnstico ms precoz.
La ecografa transvaginal permite hacer un diagnstico de embarazo de
certeza y precoz.
RECUERDA
HCG en sangre: mtodo ms precoz.
HCG en orina: mtodo ms habitual.
Eco: diagnstico de certeza.
La HCG aumenta su concentracin plasmtica en forma exponencial
durante las primeras 12 semanas de gestacin, para luego ir en paulati-
no descenso hasta fnes del embarazo. Cumple su funcin (mantener el
cuerpo lteo viable) hasta que la placenta se encuentra preparada para
producir cantidades sufcientes de progesterona, necesaria para mante-
ner el endometrio secretor (Figura 26).
Figura 26. Estimulacin hormonal del endometrio
Manual CTO 1. Edicin
16
8.5. Compatibilidad inmunolgica
La implantacin, vista desde el punto de vista inmunolgico, es la intro-
duccin de un grupo celular (el embrin) en otro (el endometrio), gen-
ticamente diferente. El embrin posee la mitad de su genoma heredado
del padre, un individuo genticamente diferente de la madre. Sera de
esperar el rechazo del embrin por parte del endometrio, hecho que no
ocurre normalmente.
Es ms, el endometrio colabora activamente con la implantacin. Se
han estudiado varias posibles respuestas al interrogante planteado
por la falta de rechazo. Todas ellas incluyen una baja capacidad an-
tignica del feto o la ocultacin de la misma por diferentes meca-
nismos.
8.6. Inactivacin
(lionizacin) del cromosoma X
La inactivacin de uno de los cromosomas X de la mujer ocurre al azar,
hacia el da 14 de vida intrauterina, y tambin se denomina lionizacin.
La manifestacin morfolgica de la lionizacin es la formacin del cor-
psculo de Barr.
8.7. Patologa de la segunda semana
Abortos
Las causas de un aborto en la segunda semana pueden ser las mis-
mas estudiadas para la primera semana, a las cuales se sumaran
las siguientes:
- Incompatibilidad inmunolgica, sera un rechazo inmunolgico
del embrin al reconocerlo como extao.
- Defciente secrecin de progesterona por parte del cuerpo lteo,
que traera como consecuencia un desequilibrio en la fase secre-
toria endometrial, necesaria para la correcta implantacin.
Embarazos ectpicos
La implantacin puede llevarse a cabo en la trompa uterina, el ova-
rio, la cavidad abdominal o el cuello uterino. Este tipo de embarazo
generalmente no llega a trmino, sobre todo los producidos en ab-
domen y trompa, que acarrean serios problemas a la madre y hacen
peligrar su vida.
El tipo ms frecuente de embarazo ectpico es el tubario (95%)
(Figura 27). Generalmente existe el antecedente de infeccin genital alta
(enfermedad pelviana infamatoria) o de endometrosis, siendo ambas
causas de cicatrizacin de la trompa o el peritoneo que la recubre, lo que
ocasiona oclusiones parciales o totales de la luz tubaria que difcultan
el traslado del embrin hacia la cavidad uterina. Este es un ejemplo de
infertilidad, ya que se forma la clula huevo pero el embarazo no llega
a trmino (diferente de esterilidad, donde no se forma clula huevo).
RECUERDA
El sitio ms frecuente de implantacin ectpica es la trompa de Falopio.
La implantacin cercana al orifcio interno del cuello uterino produce
placenta previa, que puede ocluir el canal de parto, produciendo he-
morragias durante los ltimos meses de gestacin.
Figura 27. Embarazo ectpico
Embarazos molares. Enfermedad trofoblstica gestacional
Durante la gametognesis ocurrira un fenmeno conocido como
impronta genmica, que consiste en modifcaciones en los patrones
de metilacin de los genes segn el sexo del individuo. Esto quiere
decir que las mujeres y los hombres metilan su ADN de diferentes
formas, le ponen una impronta que es propia del sexo al que per-
tenece el progenitor. La metilacin del ADN es una de las formas de
regulacin de la expresin gnica.
La mola, o embarazo molar, se caracteriza por el desarrollo excesivo
de los tejidos trofoblsticos. Aparentemente, la expresin temprana
de los genes maternos, determina el desarrollo inicial del embrio-
blasto, mientras que la de los genes paternos, regula el desarrollo
del trofoblasto.
La mola completa es diploide y carece de embrin en su interior.
Se formara por la fecundacin de un ovocito vaco (carente de
ncleo) por dos espermatozoides, o por uno que se duplica en su
interior. Al carecer de genoma metilado al estilo materno, no ini-
cia el desarrollo embrionario. Por poseer dos copias del genoma
paterno, el trofoblasto se desarrolla en exceso. Estas pacientes se
presentan clnicamente con sangrado genital en el primer trimes-
tre, y ocasionalmente eliminan un material de aspecto vesiculoso,
en racimo de uvas. El anlisis microscpico del material de aborto
muestra vellosidades coriales edematosas (degeneracin hidrp-
ca), avasculares (los vasos de las vellosidades coriales pertenecen
al embrin, que en este caso no existe). NO se aisla embrin y el
cariotipo es diploide. La secrecin de hormona HCG es marcada-
mente mayor que la esperada para la edad gestacional, ya que el
trofoblasto es excesivo. El dosaje seriado de esta hormona es de
suma importancia en el seguimiento de estas pacientes, ya que un
aumento de sus valores puede indicar la recurrencia de la enfer-
medad.
RECUERDA
La mola completa es diploide (46 XY) y carece de embrin en su interior.
EMBRIOLOGA 9
17
La mola incompleta o parcial, en cambio, posee tanto tejidos
embrionarios como extra-embrionarios, stos ltimos en exceso.
Se formara merced a la unin de un ovocito normal (haploide), fe-
cundado por dos espermatozoides normales o por uno diploide. A
consecuencia de ello se forma un embarazo molar con un embrin
triploide, no viable a largo plazo. La presencia de genoma materno
determina el inicio del desarrollo embrionario, y el genoma pater-
no (doble) es responsable del tejido trofoblstico excesivo. Clnica-
mente estas pacientes presentan sangrado genital, habitualmente
en etapas ms avanzadas de la gestacin. El anlisis del material
eliminado del tero puede no sugerir macroscpicamente la pre-
sencia de esta patologa, ya que algunas de las vellosidades coriales
sern avasculares, edematosas (degeneracin hidrpica), y otras
sern normales. Si se aisla embrin, este presenta malformaciones
congnitas severas. El seguimiento tambin se realiza con dosajes
seriados de HCG.
RECUERDA
La mola parcial es triploide (46 XXY) y presenta tejido embrionario y/o
amnios.
La importancia de identifcar y controlar estas patologas radica en la
posibilidad de persistencia de la enfermedad (enfermedad trofoblstica
persistente) o el eventual desarrollo de un tumor maligno, el coriocarci-
noma, que en el 1-2% de los casos tiene como antecedente un embarazo
molar. El riesgo de enfermedad trofoblstica persistente es mayor con la
mola completa.
9. Tercera semana. Gastrulacin
El evento principal de la tercera semana de gestacin es la gastrulacin,
proceso mediante el cual se genera un embrin trilaminar, formado
por las tres hojas (endodermo, mesodermo, ectodermo) que darn ori-
gen a todos los tejidos embrionarios.
El mecanismo biolgico del desarrollo ms caracterstico de este perodo
es la haptotaxis, que es la migracin celular siguiendo un gradiente
qumico presente en la matriz extracelular (MEC).
9.1. Anatoma del embrin
de 14 das-territorios presuntivos
Nuestro embrin de fnes de segunda semana era bilaminar (epiblasto
e hipoblasto), aunque en realidad todos los tejidos intraembrionarios (y
algunos extraembrionarios) van a derivar de una sola de sus lminas, el
epiblasto. La haptotaxis es entonces el mecanismo biolgico ms rele-
vante, ya que las clulas debern migrar, desde su ubicacin en el epi-
blasto, hasta sus posiciones defnitivas.
RECUERDA
El embrin de la segunda semana es bilaminar, pero slo el epiblasto va a
dar derivados intraembrionarios.
El da 15 del desarrollo embrinario aparece un surco a lo largo de la lnea
media caudal del disco embrionario bilaminar. Progresivamente va ha-
cindose ms alargada y profundo (Tabla 4).
El da 16 de desarrollo embrionario, en la parte craneal de dicho surco, se
forma una depresin rodeada de un reborde de epiblasto.
Lnea primitiva
Surco = surco primitivo
Depresin = fosita primitiva
Reborde = ndulo primitivo o ndulo de Hensen
Tabla 4. Territorios embrionarios
RECUERDA
La lnea primitiva aparece al comienzo de la tercera semana.
Entonces podemos decir que, desde principios de la tercera semana
(da 14-15), pueden reconocerse en el embrin la asimetra derecha-
izquierda y cfalo-caudal.
Por ltimo, pueden observarse dos reas sobreelevadas a nivel del epi-
blasto, que corresponden en realidad a zonas en las que el epi e hipoblas-
to (y luego el endodermo) se encuentran frmemente adheridos. Una de
ellas es la membrana bucofarngea u orofarngea, a nivel ceflico, y la
otra es la membrana cloacal, a nivel caudal.
A nivel del epiblasto, se encuentran representados numerosos tejidos,
tanto intra como extraembrionarios. Cada sector del epiblasto que repre-
senta un tejido es un territorio presuntivo (TP). Se han identifcado los
siguientes territorios presuntivos:
TP del mesodermo extraembrionario.
TP del endodermo extraembrionario o hipoblasto.
TP del endodermo (intraembrionario).
TP del mesodermo intraembrionario.
TP de la notocorda.
TP del ectodermo general.
TP del ectodermo neural.
Un territorio presuntivo es un rea del epiblasto cuyas clulas migran
siguiendo un destino evolutivo, que est defnido por el lugar que esas
clulas ocupan en el espacio y NO por las caractersticas intrnsecas de
las mismas.
Las clulas de un TP tienen igual potencialidad evolutiva que las de otro
TP, lo importante justamente no son las clulas, sino el sustrato, la matriz,
el medio ambiente en el que las clulas se encuentran que determinarn
que esa clula se diferencie hacia un determinado rgano o sistema.
RECUERDA
Los territorios presuntivos se ubican en el epiblasto.
Las clulas de un territorio presuntivo se determinan y diferencian en un
sentido evolutivo despus de haber migrado. El nico TP que se encuen-
tra determinado desde un comienzo es el de la notocorda, es decir que
si movemos las clulas de este TP hacia el TP del ectodermo, se generar
Manual CTO 1. Edicin
18
a ese nivel otra notocorda. Por eso la notocorda (que como TP est repre-
sentada por el nodo de Hensen) es considerada la organizadora.
RECUERDA
El nico territorio presuntivo que se encuentra determinado antes de su
migracin es el de la notocorda
9.2. Migracin de TP extraembrionario
La migracin del TP del mesodermo extraembrionario se produjo a mediados
de la segunda semana, cuando las clulas, que ocupaban un rea en la parte
caudal del epiblasto, se desplazaron fuera de ste para ubicarse por dentro
del citotrofoblasto y por fuera de la cavidad amnitica y el saco vitelino.
La migracin del TP del endodermo extraembrionario (o hipoblasto) tam-
bin se produjo en la segunda semana. Las clulas de este TP convergen en
la estra primitiva y luego migran en sentido ventral (hacia el saco vitelino),
emplazando el hipoblasto antiguo, que se encontraba formando el techo
de esta cavidad. A medida que el hipoblasto es desplazado de su ubicacin,
sus clulas se integran a las paredes del saco vitelino. Despus de la migra-
cin de los TP extraembrionarios, al epiblasto lo llamamos embrioblasto,
ya que la totalidad de sus clulas darn derivados intraembrionarios.
9.3. Migracin de TP intraembrionario
9.3.1. TP del endodermo
El da 16, las clulas del epiblasto prximas a la lnea primitiva proliferan
y pierden las uniones intercelulares. Estas clulas se aplanan y emiten
unas prolongaciones llamadas pseudpodos que les permiten emigrar
a travs de la lnea primitiva y posicionarse en el espacio existente entre
el epiblasto y el hipoblasto. Este proceso de involucin y penetracin se
llama gastrulacin. Algunas clulas del epiblasto que emigran a travs de
la lnea primitiva sustituyen la capa de clulas del hipoblasto, sustituyn-
dolo y formando una capa de clulas llamada ahora endodermo.
As, queda formado el embrin por dos hojas (embrin bilaminar): endo-
dermo (ventral) y epiblasto (dorsal).
Recordemos que existen dos zonas donde endodermo y epiblasto se en-
cuentran fuertemente adheridos, la membrana bucofarngea y la mem-
brana cloacal, en los extremos ceflico y caudal del disco embrionario
respectivamente. Al componente endodrmico de la membrana buco-
farngea se lo conoce tambin como placa procordal (o precordal).
9.3.2. TP de la notocorda-mesodermo precordal
El TP de la notocorda se ve representado por el ndulo de Hensen o nodo
primitivo (Figura 28). Para formarlo, las clulas convergen en la lnea me-
dia. Luego invaginan y comienzan a migrar en sentido ceflico (elongan).
En su interior, la fosita tambin crece en sentido ceflico.
RECUERDA
Los movimientos para formar la notocorda son: convergencia, invaginacin
y elongacin. La notocorda se forma a partir de las clulas que penetran a
travs del ndulo primitivo.
Como consecuencia, entre el epiblasto y el endodermo, queda consti-
tuido el conducto notocordal, que contina comunicado con la cavidad
amnitica por el orifcio original de la fosita primitiva. A medida que la
notocorda elonga hacia ceflico, la lnea primitiva retrocede hacia caudal,
Figura 28. Formacin de la notocorda
EMBRIOLOGA 9
19
vale decir que la fosita primitiva tambin lo hace, elongando de esta for-
ma la notocorda tambin en sentido caudal.
Las clulas del endodermo y la pared ventral del conducto notocordal de-
generan temporariamente y se comunican el saco vitelino y la cavidad
amnitica. Esta comunicacin se conoce como conducto neuroentri-
co y el tejido notocordal remanente adopta la forma de placa.
RECUERDA
El conducto neuroentrico comunica temporalmente la cavidad amnitica
con el saco vitelino denitivo
El endodermo posteriormente se regenera, quedando la notocorda, aho-
ra un cordn macizo, atrapada entre el ectodermo y el endodermo.
El TP de la notocorda migra en sentido ceflico hasta el mesodermo pre-
cordal, que se ubica en la parte ceflica del disco embrionario, justo caudal
a la membrana bucofarngea (es decir, que este tejido migr por el mis-
mo trayecto que el TP de la notocorda pero previo a sta). El mesodermo
precordal tiene importancia en la induccin del desarrollo del cerebro
anterior. No debe confundirse con la placa precordal (o procordal), que
corresponde al endodermo de la membrana bucofarngea.
9.3.3. TP del mesodermo intraembrionario
Las clulas que forman este TP inicialmente convergen para formar la lnea
primitiva, luego invaginan en la lnea media del disco, y divergen, forman-
do dos alas de mesodermo. Estas alas crecen hacia el extremo ceflico del
disco (elongan), igual a como lo hicieron las clulas de la notocorda.
Los movimientos de elongacin de la notocorda y del mesodermo intra-
embrionario son simultneos.
Sin embargo, las clulas del mesodermo intraembrionario no son frena-
das por el mesodermo precordal (que a su vez fue frenada por la placa
procordal de la membrana bucofarngea), sino que pasan por sus costa-
dos, hasta el extremo ceflico del disco, donde reconvergen (es decir, se
acercan a la lnea media hasta fusionarse un ala con la opuesta).
Los movimientos del TP del mesodermo IE son:
Convergencia.
Invaginacin.
Divergencia.
Elongacin hasta el extremo ceflico.
Reconvergencia (por delante de la membrana bucofarngea).
Conclusiones:
El TP de la notocorda es el ndulo de Hensen.
Numerosos TP (el del endodermo, el del mesodermo IE) migran des-
de su localizacin inicial en el epiblasto hasta su destino final, pasan-
do por la lnea o estra primitiva.
Simultnea a la migracin de la notocorda por la lnea media, se pro-
duce la migracin del mesodermo IE, por los laterales del disco.
A medida que las clulas migran, la lnea primitiva retrocede, hasta des-
aparecer, hacia principios de la cuarta semana. El ltimo tejido formado
ser el mesodermo correspondiente a la regin caudal del cuerpo, cono-
cido como eminencia caudal. Es notable cmo la porcin caudal del em-
brin muestra esta importante actividad proliferativa hasta fnes de la
tercera semana, migrando luego las clulas desde las porciones ms cau-
dales hacia las ceflicas. La porcin ceflica, en cambio, muestra un nota-
ble crecimiento (a expensas de las clulas migrantes) y mayor grado de
diferenciacin. Estos hechos que se inician posteriores a la gastrulacin
sern una constante en el desarrollo embrionario y fetal, observndose
siempre mayor tamao y complejidad a nivel ceflico, los que se trasladan
en el desarrollo en sentido caudal (un ejemplo de esto es la formacin de
los miembros: los superiores aparecen antes que los inferiores) (Figura 29).
Figura 29. Migracin celular del epiblasto hasta
su futura localizacin defnitiva
Evolucin del mesodermo intraembrionario (Figura 30).
El mesodermo IE presenta diferentes regiones, que se distinguen ya des-
de su migracin (de hecho ciertos textos describen en la lnea primitiva,
un sector destinado al mesodermo lateral, otro al intermedio, etc., es de-
cir que estas regiones del mesodermo podran considerarse diferentes al
TP en el epiblasto). Estas regiones son:
La placa cardiognica, con forma de herradura, en el extremo cef-
lico del disco.
El mesodermo branquial, entre las ramas de la herradura antes
mencionada.
Los somitas o mesodermo paraaxial, barras macizas pares de meso-
dermo ubicadas en la parte dorsal del embrin, de las cuales se origi-
narn las vrtebras, costillas y msculos del tronco.
El mesodermo correspondiente a los gononefrtomos o meso-
dermo intermedio, que son dos cordones macizos ubicados por fue-
ra de los somitas, y que darn origen a los aparatos genital y urinario.
El mesodermo lateral, por fuera del anterior.
El mesodermo caudal.
A nivel de la placa cardiognica y del mesodermo lateral, y como am-
bas partes se encuentran divididas en las hojas visceral y parietal, que-
da defnida una cavidad, el celoma intraembrionario. Decimos que a
este nivel el mesodermo se delamina (en dos lminas u hojas, como
sucedi en la segunda semana con el mesodermo extraembrionario),
y se produce la celomizacin (aparicin de una cavidad) a nivel intra-
embrionario.
Manual CTO 1. Edicin
20
9.3.4. TP del ectodermo general
El movimiento que defne la formacin del ectodermo general es la epi-
bolia, que signifca proliferacin y migracin de clulas del epiblasto
para rellenar los espacios vacos dejados por las clulas que migraron
desde otros TP.
9.3.5. TP del ectodermo neural (Figura 31)
Bajo la induccin del mesodermo precordal y la notocorda (y mien-
tras sta se genera), las clulas del ectodermo que se encuentran por
encima de ella aumentan de altura, volvindose cilndricas, formando
de esta manera la placa neural. A medida que la notocorda se alarga,
la placa neural se ensancha y se extiende en sentido craneal hasta la
membrana bucofarngea. Finalmente, se prolonga ms all de la no-
tocorda.
Tenemos entonces un TP del ectodermo neural, esta placa, que pos-
teriormente, por contraccin del citoesqueleto de su porcin apical,
formar un surco, el surco neural. Al acercarse ambos extremos (o
labios) de este surco, el neuroectodermo comienza a transformarse
en un tubo, el tubo neural, que toma independencia del ectodermo
general y se ubica ventral a ste y dorsal a la notocorda. Al proceso de
formacin de la placa, surco y tubo neural se le llama en su conjunto
neurulacin.
Al formarse el tubo neural, dos cordones de clulas se desprenden de sus
laterales, formando las crestas neurales, que tienen entre sus derivados
al sistema nervioso perifrico y numerosas estructuras no neurales.
Figura 31. Neurulacin y formacin de la cresta neural
Antes del cierre completo del tubo neural, que ocurre hacia fnes de la cuar-
ta semana, dos orifcios (uno craneal y otro caudal) comunican su luz con la
cavidad amnitica. Estos orifcios toman el nombre de neuroporos.
RECUERDA
La placa neural se forma por induccin de la notocorda. Los neuroporos
comunican el interior del tubo neural con la cavidad amnitica.
Figura 30. Partes del mesodermo intraembrionario
EMBRIOLOGA 9
21
9.4. Evolucin de los anejos embrionarios
Saco vitelino: en las paredes del saco vitelino, que, al igual que a fnes de
la segunda semana, se encuentra formado por endodermo y por la hoja
visceral del mesodermo extraembrionario, van a ocurrir eventos de gran
importancia.
En el mesodermo extraembrionario se van a formar los primeros va-
sos sanguneos (Figura 32). El proceso puede resumirse como sigue:
Se forman acmulos de clulas denominadas angioblastos. Son bo-
las pequeas, macizas, llamadas islotes de Wolff y Pander.
Los angioblastos de la periferia se aplanan y forman una estruc-
tura similar a una pared (la pared vascular), mientras que las que
quedan en el centro forman las clulas sanguneas primitivas o
megaloblastos.
Este evento ocurre en varios lugares a la vez (siempre dentro del meso-
dermo del saco vitelino), por lo que estas estructuras vasculares se fusio-
nan unas a otras, hasta dejar formados los vasos sanguneos tal cual los
conocemos. Si bien en un principio el fenmeno es exclusivo del saco
vitelino, poco despus se expande al resto del mesodermo, tanto ex-
tra como intraembrionario, quedando formados los vasos primitivos del
embrin y los de las vellosidades coriales.
RECUERDA
Los vasos sanguneos y las clulas sanguneas son siempre de origen
mesodrmico.
Figura 32. Formacin de vasos sanguneos
Sobre las clulas germinales podemos decir que aparecen durante la ter-
cera semana, en el endodermo que tapiza al saco vitelino, y que luego
migrarn hacia sus lugares de destino (las gnadas primitivas). El origen
de estas clulas NO es el saco vitelino como muchos textos describen.
En realidad, y este concepto es relativamente nuevo, si rastreamos el ori-
gen de estas clulas, podemos ya desde las primeras blastmeras gene-
radas en la segmentacin, identifcar un grupo que dar como derivado
a las clulas germinales. Este grupo luego migrara hacia el epiblasto y
posteriormente al saco vitelino. Globalmente, la mayora de los autores
considera que en el epiblasto se observa un TP que corresponde a las
clulas germinales primitivas por lo que podemos tomar el epiblasto
como el origen (si bien sabemos que en realidad estn determinadas
desde mucho antes).
Vellosidades coriales (Figura 33).
Las vellosidades de fnes de la segunda semana eran cordones macizos
nacidos del citotrofoblasto rodeadas por sincitiotrofoblasto.
Figura 33. Vellosidades coriales.
Durante la tercera semana, estas vellosidades van a evolucionar. Primero,
se va a introducir en ellas mesodermo extraembrionario (hoja parietal),
por lo que ahora se van a llamar vellosidades secundarias (Figura 34).
Figura 34. Vellosidades secundarias
Dentro de este mesodermo comienzan a formarse vasos sanguneos, que-
dando constituidas las vellosidades terciarias o defnitivas (Figura 35).
Manual CTO 1. Edicin
22
Figura 35. Vellosidades terciarias
El citotrofoblasto de la punta de algunas vellosidades forma primero
unos botones (engrosamientos) citotrofoblsticos, cuyas clulas comien-
zan a migrar hacia la periferia del sincitiotrofoblasto, para formar la coraza
citotrofoblstica. La coraza es entonces una capa de trofoblasto interme-
dio que se interpone entre el endometrio y el sincitiotrofoblasto, y cuya
funcin es la de evitar que ste siga penetrando el tejido endometrial. A
estas vellosidades que estn conectadas con la coraza por citotrofoblasto
las llamamos vellosidades de anclaje.
9.5. Evolucin del sistema circulatorio primitivo
El aparato circulatorio del embrin de 21 das est formado por los va-
sos vitelinos (que irrigan las paredes del saco vitelino), las aortas pri-
mitivas (que transitan por el mesodermo intraembrionario) y los vasos
umbilicales (que circulan por el pedculo de fjacin y las vellosidades)
(Figura 36).
Figura 36. Aparato circulatorio embrionario
Los tubos cardacos primitivos (ubicados en la hoja visceral de la placa
cardiognica) comienzan a latir el da 21-22 de vida embrionaria, bom-
beando sangre a las arterias aortas. La sangre circula luego hacia las
arterias vitelinas y unbilicales (ramas de las aortas). Las art. umbilicales
transitan por el pedculo de fjacin, llevando la sangre a los capilares de
las vellosidades coriales, donde se produce el intercambio gaseoso (con
sangre materna presente en la red lacunar).
Las venas umbilicales, con sangre rica en oxgeno, regresan por el pe-
dculo hacia ambos tubos cardacos. Las venas vitelinas traen sangre
carboxigenada del saco, por el mismo recorrido que las arterias hom-
nimas.
Mientras que nuestro embrin de segunda semana reciba los nutrientes
por difusin desde las lagunas en el sinciciotrofoblasto, el embrin de
tercera semana, que es ms grande y necesita ms nutrientes, lo hace a
partir del intercambio que se realiza a nivel de las vellosidades terciarias.
Los nutrientes, que viajan por la sangre, se distribuyen gracias a la presen-
cia de los vasos embrionarios.
9.6. Tercera y cuarta determinaciones
La tercera determinacin est dada por el efecto que ejerce el endoder-
mo sobre el epiblasto para la generacin del mesodermo.
La cuarta determinacin consiste en la induccin de la notocorda sobre el
ectodermo general para formar el ectodermo neural.
Tanto en la tercera como cuarta determinaciones, el estmulo determi-
nante es una poblacin celular (y no una sustancia soluble del medio
ambiente, como sucedi en la primera y segunda determinaciones). En
este caso hablaremos entonces de dos poblaciones celulares:
Una poblacin celular determinante, que genera el estmulo.
Una poblacin celular competente, capaz de responder a ste.
9.7. Patologa
Neurulacin
Las alteraciones de la neurulacin originan anomalas graves del encfalo y
la mdula espinal. La meroanencefalia (mero = parcial, anencefalia = ausen-
cia de cerebro) es el defecto ms grave y tambin la anomala ms comn.
El dfcit de cido flico (un elemento esencial que debe formar parte de
la dieta) se encuentra asociado a mayor incidencia de defectos del cierre
del tubo neural.
Niveles aumentados de alfa-fetoproteina en suero materno o en lquido
amnitico se relacionan con defectos del tubo neural y otras anomalas
(rin poliqustico, onfalocele, sndrome de Turner con higroma qustico,
atresia duodenal).
10. Cuarta semana
Durante la cuarta semana se forman los esbozos de la mayora de los
rganos importantes. El disco embrionario sufre una serie de plega-
mientos debido a que su crecimiento diferencial le obliga a plegarse
EMBRIOLOGA 9
23
y pasa a formarse una estructura tridimensional vertebrada. El primer
plegamiento se realiza en el eje longitudinal convirtiendo al embrin
en una estructura convexa. Este plegamiento comienza a nivel cefli-
co y culmina con el plegamiento caudal que hace que el pedculo de
fjacin se acerque al saco vitelino (Figura 37).
Figura 37. Plegamientos embrionarios
El segundo plegamiento consiste en la aproximacin y posterior fusin
de los bordes laterales a lo largo de la lnea media ventral. La fusin del
endodermo en la lnea media produce el primordio del intestino que
ahora se divide en intestino anterior, intestino medio (unido al saco vite-
lino por el conducto vitelino) e intestino posterior.
Del intestino anterior derivarn el estmago, el duodeno, el hgado, el
pncreas y la vescula biliar. El bazo no deriva del endodermo, sino del
mesogastrio dorsal. Del intestino posterior derivan el recto y el seno uro-
genital.
La falta de cierre completo del ombligo produce el onfalocele. El con-
ducto vitelino o conducto onfalomesentrico regresa en condiciones
normales entre la quinta y la octava semana, pero en un 2% de los recin
nacidos vivos persiste el conducto onfalomesentrico formando general-
mente un divertculo intestinal llamado divertculo de Meckel.
RECUERDA
El divertculo de Meckel es la persistencia del conducto onfalomesentrico.
El plegamiento hace que la cavidad celmica intraembrionaria pase a ser
una cavidad cerrada. Del mesodermo lateral se formarn las serosas, la
cavidad pericrdica, pleural y peritoneal.
Sucesivamente de craneal a caudal han ido apareciendo los somitas, que
son derivados del mesodermo paraaxial (Figura 38). En la regin cervical
los somitas pueden dividirse en miotomos (que darn lugar a los ms-
culos de la espalda y pared antero-lateral del cuerpo), dermatomos (que
formarn la dermis del cuero cabelludo, cuello y tronco) y esclerotomos
(que formarn el cuerpo y arco vertebrales y contribuirn a la formacin
de la base del crneo). Las clulas de los esclerotomos rodean a la noto-
croda y al tubo neural.
La formacin del tubo neural comienza el vigsimo segundo da a ni-
vel de los primeros cinco somitas. El cierre del tubo neural progresa en
ambas direcciones y concluye con el cierre de los neuroporos ceflico y
caudal. Durante la cuarta semana ya estn formados los esbozos de las
principales partes del encfalo y la mdula espinal. La porcin ms cau-
dal del tubo neural se forma por neurulacin secundaria de la prominen-
cia caudal.
De los pliegues neurales surgen las clulas de la cresta neural que emi-
gran a diferentes localizaciones dando lugar a diferentes estructuras
como ganglios sensitivos del sistema nervioso perifrico (SNP), melano-
citos, zonas de la mdula suprarrenal, etc.
El da vigsimo segundo se forma el primordio respiratorio y las yemas
pulmonares. Continuarn desarrollndose pero no madurarn hasta la
aparicin de surfactante antes del nacimiento.
RECUERDA
La causa ms frecuente de hipoplasia pulmonar es la hernia diafragmtica
congnita.
El septum transversum separa parcialmente la cavidad torcica de la ab-
dominal. El cordn umbilical se forma cuando el pedculo de fjacin y el
conducto vitelino se renen por la expansin del amnios. La formacin
del saco pericrdico y el diafragma entre las semanas quinta y sptima
subdivide la cavidad celmica en cuatro cavidades.
El diafragma defnitivo posee derivados de cuatro estructuras embriona-
rias: septum transversum, membranas pleuroperioneales, mesodermo
paraaxial de la pared del cuerpo y del mesnquima esofgico.
Manual CTO 1. Edicin
24
11. Desarrollo del corazn
En el disco embrionario ya exista una zona llamada regin cardiognica
que es donde se va a originar el futuro corazn. Por diferentes seales ad-
yacentes a esta zona, se estimula la formacin de dos tubos sanguneos pa-
ralelos llamados tubos endocrdicos laterales. Los diferentes plegamientos
que va sufriendo el embrin en la cuarta semana hacen que estos tubos
endocrdicos laterales se vayan aproximando y acaben fusionndose for-
mando el tubo cardaco primitivo.
Al mismo tiempo se van formando el par de aortas dorsales del sistema
circulatorio primitivo que darn lugar a la futura aorta.
Este tubo cardaco primitivo sufre una serie de movimientos progresivos
(giro, tabicacin, pliegues y remodelacin) desde la quinta a la octava
Figura 38. Los somitas y sus partes
EMBRIOLOGA 9
25
semana que darn lugar fnalmente al corazn defnitivo con cuatro ca-
vidades y sus vlvulas.
La tabicacin de las aurculas y la divisin del canal auriculoventricular
comienzan en la cuarta semana. A fnales de la cuarta semana el sep-
tum primum comienza a crecer hacia abajo desde la raz auricular, es un
septo fno y membranoso. El orifcio que hay entre las dos aurculas que
progresivamente va hacindose ms pequeo por el crecimiento del sep-
tum primum se denomina ostium primum. En el septum primum se van
produciendo unas perforaciones en la parte superior que coalescen para
formar el ostium secundum. Al mismo tiempo comienza a formarse un
nuevo septo grueso y muscular al lado del septum primum que se llama
septum secundum. El septum secundum tambin tiene un orifcio que se
denomina agujero oval. Durante el desarrollo fetal la sangre de la aurcu-
la derecha atravesar el orifcio oval y el ostium secundum para pasar a la
aurcula izquierda. Estos dos orifcios se cerrarn en el nacimiento.
La tabicacin de los ventrculos est coordinada con la formacin de las
vlvulas auriculoventriculares. (Figuras 39, 40 y 41)
Figura 39. Tabicacin de las aurculas
Figura 41. Tabicacin de los ventrculos
12. Sistema excretor-genital
Los dos sistemas se desarrollan en paralelo y derivan del mesodermo in-
termedio. Es un sistema par (a la derecha y a la izquierda) y con disposi-
cin metamrica.
El sistema excretor empieza a diferenciarse en la cuarta semana. A
nivel cervical est el sistema pronfrico que desaparece a fnales de
la cuarta semana y a nivel torcico est el sistema mesonfrico que
desaparecer parcialmente. Se forma un tercer sistema a nivel lum-
Figura 40. Formacin del corazn
Manual CTO 1. Edicin
26
bar baja y sacra: el sistema metanfrico, que dar lugar al rin y al
sistema excretor defnitivo. El conjunto de los tres sistemas a lo largo
del embrin que proceden del mesodermo intermedio se denomina
cordn nefrgeno.
En el mesodermo se diferencian unos tubos que desembocan en el
seno urogenital, son los conductos mesonfricos o de Wolf. En la
quinta semana estos conductos se introducen en forma de yema urete-
ral en el metanefros (que sern los riones defnitivos). La yema se en-
sancha formando la pelvis renal y se subdivide dando lugar a los clices
mayores y menores y los conductos colectores. De la yema tambin se
formarn los urteres.
Del metanefros derivarn las nefronas, que son unidades del sistema ex-
cretor compuestas por la cpsula de Bowman, el tubo contorneado proxi-
mal, el asa de Henle y el tubo contorneado distal.
El alantoides degenera formando el ligamento uraco, que sujeta la vejiga a
la pared del cuerpo. La vejiga y la uretra se forman a partir del endodermo.
El trgono vesical (parte posterior de la vejiga) es derivado mesodrmico.
El mesodermo que hay en las crestas genitales es el que dar lugar a las
gnadas. Las clulas germinales primordiales se colocan en la cresta geni-
tal en forma de cordones sexuales primarios. Hasta la sptima semana esta
estructura es similar en los varones y en las mujeres (perodo indiferente).
En la cuarta-quinta semana se van formando otros conductos paralelos
a los conductos mesonfricos o de Wolf que se llaman conductos para-
mesonfricos o de Mller.
Diferenciacin masculina: en el interior de la gnada (parte medular de
la gnada) se diferencian muchos cordones medulares. Estos cordones
medulares formarn los tubos seminferos y las clulas de Sertoli. El resto
del mesodermo de la gnada que est entre los cordones dar lugar a las
clulas de Leydig. La meiosis no comenzar hasta la pubertad en el varn.
Las clulas de Leydig liberan testosterona. Las clulas de Sertoli secre-
tan hormona antimlleriana que hace que desaparezca el conducto de
Mller. El conducto mesonfrico dar lugar al conducto deferente que
se enrolla en la parte proximal formando el epiddimo. La cubierta que
cubre el testculo, los conductos eferentes y el epiddimo es la tnica al-
bugnea que es un derivado del epitelio celmico. El tubrculo genital
dar lugar al glande.
La prstata es un derivado del endodermo. La uretra es un derivado en-
dodrmico excepto su porcin fnal que es ectodrmica.
Diferenciacin femenina: los cordones invaden la zona externa de la
gnada (corteza) formando cordones corticales. Estos cordones formarn
las clulas foliculares. Las ovogonias comienzan la meiosis.
En la mujer no se secreta testosterona por lo que los conductos y t-
bulos mesonfricos desaparecen (por supuesto una vez ya formada la
yema ureteral). Como no existen clulas de Sertoli, no se secreta hor-
mona antimlleriana y los conductos de Mller permanecen. La parte
proximal de los conductos de Mller dar lugar a las trompas de Falopio
y la porcin donde se unen en la lnea media formar el tero, el con-
ducto utero-vaginal y la partems craneal (1/3 superior) de la vagina. La
porcin inferior de la vagina (2/3 inferiores) se forma a partir del endo-
dermo (seno urogenital). El tubrculo genital dar lugar al cltoris.
Las gnadas de ambos sexos que se haban formado a nivel torcico-
lumbar tienen que migrar hacia caudal para adoptar sus posiciones de-
fnitivas.
13. Placenta
La placenta se forma de la unin de un componente fetal, el corion fron-
doso (con vellosidades terciarias), y un componente materno, la decidua
basal. La funcin ms importante de la placenta es el intercambio de sus-
tancias.
La sangre materna (con oxgeno) entra por la vena umbilical y llega al
corazn del embrin, pasa a la arteria aorta que la distribuye y llega a
la arteria umbilical ya sin oxgeno, va a las vellosidades para realizar el
intercambio gaseoso.
Se habla de placenta a partir del cuarto mes de embarazo. Del cuarto
al decimo mes sufre un perodo de maduracin que consiste en au-
mentar la complejidad de las vellosidades coriales y en adelgazar la
barrera placentaria.
Las funciones de la placenta son:
Barrera: evita la comunicacin directa entre circulacin materna y fe-
tal y protege al feto de ciertas sustancias.
Endocrina: segrega progesterona.
Respiracin, nutricin y excrecin del feto.
14. Teratogenia
Recordemos frmacos que producen alteraciones en el feto si se adminis-
tran durante el embarazo:
Quinolonas: alteraciones en el cartlago de crecimiento.
Tetraciclinas: manchas en los dientes.
Sulfamidas: kernicterus.
Aminoglucsidos: nefro y ototoxicidad.
Fenobarbital: alteraciones hepticas.
Bibliografa
Embriologa clnica; K. L. Moore, T. V. N. Persaud; 7. ed.; Elsevier;
2004.
Embriologa humana; W. J. Larsen; 3.
a
ed.; Elsevier; 2003.
Embriologa mdica; J. Hib; 8.
a
ed.; Editorial Clareo; 2006.
Embriologa mdica con orientacin clnica; T. W. Sadler; 8.
a
ed.; Edito-
rial Mdica Panamericana; 2001.
Gentica humana: fundamentos y aplicaciones en medicina; A. J. Sola-
ri; 1. ed.; Editorial Mdica Panamericana; 1996.
27
DIGESTIVO 8
Conceptos clave
El perodo preembrionario comprende las tres primeras semanas de vida intrauterina.
El perodo embrionario abarca de la cuarta a la octava semana.
El perodo fetal comprende de la novena semana hasta el nacimiento.
El espermatozoide y el ovocito son clulas haploides.
La formacin de gametos se realiza mediante la divisin meitica.
Los ovocitos I estn detenidos en la profase I (dictiotene) hasta la pubertad.
Los ovocitos slo completarn la segunda meiosis si hay fecundacin.
La espermatognesis comienza en la pubertad y las dos divisiones meiticas se realizan secuencialmente.
El ciclo sexual femenino normal dura 28 das y tiene dos fases: la primera proliferativa dependiente de estr-
genos y la segunda secretora dependiente de la progesterona.
El pico de LH es el responsable de la ovulacin.
El ovocito es ovocito primario en profase de la 1. meiosis hasta la pubertas. Con la ovulacin, se completa la
1 meiosis y pasa a ser ovocito secundario hasta la fecundacin, que estimula la segunda divisin meitica.
La membrana pelcida debe mantenerse hasta momentos antes de la implantacin.
El espermatozoide madura en el tracto genital masculino pero se capacita en el femenino.
La fecundacin se produce en el tercio distal de la trompa de Falopio.
Se forma una clula diploide a partir de dos clulas haploides.
El bloqueo de la poliespermia impide que un mismo ovocito sea fecundado por ms espermatozoides.
El sexo depende del cromosoma sexual que lleve el espermatozoide.
La segmentacin hace que el cigoto se divida sin aumentar de tamao.
La implantacin se produce el 5-6 da en forma de blastocisto.
Del trofoblasto deriva el citotrofoblasto y el sincitiotrofoblasto.
Del embrioblasto derivan el epiblasto, el hipoblasto y el amnios.
El embrin bilaminar pasa a ser trilaminar al introducirse mesodermo entre el ectodermo y el endodermo.
La notocorda se forma a partir de las clulas que penetran a travs del ndulo primitivo.
Niveles elevados de alfa-fetoprotena en suero materno o en lquido amnitico indican defectos en el tubo
neural.
El disco embrionario sufre plegamientos.
Se forman los esbozos de la mayora de los rganos.
EL divertculo de Meckel es la persistencia del conducto onfalomesentrico.
Las clulas de la cresta neural migran a diferentes localizaciones.
Para formarse el corazn tiene que sufrir una serie de pliegues, remodelacin y tabicaciones.
El sistema excretor y genital derivan del mesodermo intermedio.
Los conductos de Wollf darn lugar a urter y la pelvis renal en ambos sexos.
Los conductos de Mller formarn en la mujer las trompas de Falopio, el tero, el conducto tero-vaginal y
el tercio superior de la vagina.
S-ar putea să vă placă și
- Anatomía & estiramientos para el embarazo y el posparto (Color): Más de 25 vídeos tutorialesDe la EverandAnatomía & estiramientos para el embarazo y el posparto (Color): Más de 25 vídeos tutorialesEvaluare: 5 din 5 stele5/5 (1)
- 01 Anatomia Desgloses 2014 WebDocument4 pagini01 Anatomia Desgloses 2014 WebDawin Tejada RodriguezÎncă nu există evaluări
- Ejemplo Preguntas Mir - PediatriaDocument2 paginiEjemplo Preguntas Mir - PediatriaLigia Zabala MoncayoÎncă nu există evaluări
- Pediatría PDFDocument12 paginiPediatría PDFChepita Pineda100% (1)
- CTO - HistologiaDocument33 paginiCTO - HistologiaRafael Torres50% (2)
- Comentarios Simulacro 1 - 2016Document86 paginiComentarios Simulacro 1 - 2016Jennifer GutiérrezÎncă nu există evaluări
- RMD 01 1819 Desgloses HMDocument6 paginiRMD 01 1819 Desgloses HMClaudia Familia100% (1)
- Examen ENURM-2023-CDocument14 paginiExamen ENURM-2023-CDra. Lady Laura Silverio100% (1)
- Desglose Actualizado CirugiaDocument41 paginiDesglose Actualizado Cirugiayols12100% (1)
- CTO 8° Edición DIGESTIVODocument271 paginiCTO 8° Edición DIGESTIVOTania Alva89% (46)
- Mir 11 2122 Preguntastestdeclase DGDocument7 paginiMir 11 2122 Preguntastestdeclase DGВероника МессерÎncă nu există evaluări
- Desgloses Cardiología CTODocument10 paginiDesgloses Cardiología CTOFranklinAntonioAcevedoDiling100% (1)
- Repaso EnurmDocument12 paginiRepaso EnurmCesarEliasÎncă nu există evaluări
- Desglose Nefrologia CtoDocument10 paginiDesglose Nefrologia CtoDanny Garcia Ramos100% (1)
- RMD 01 1819 Desgloses NRDocument4 paginiRMD 01 1819 Desgloses NRClaudia FamiliaÎncă nu există evaluări
- Plan de Estudio NefrologiaDocument6 paginiPlan de Estudio NefrologiaJames Cojab SacalÎncă nu există evaluări
- ME Desg2017 Index PDFDocument5 paginiME Desg2017 Index PDFEle PG0% (1)
- Desgloses Digestivo CTODocument7 paginiDesgloses Digestivo CTOFranklinAntonioAcevedoDilingÎncă nu există evaluări
- Desglose Pediatria 2020Document29 paginiDesglose Pediatria 2020JAZMINÎncă nu există evaluări
- XIV Curso de Residentes de Medicina Interna PDFDocument218 paginiXIV Curso de Residentes de Medicina Interna PDFsoyfenix80% (5)
- Enurm 2022 B, Resp - BibliogDocument33 paginiEnurm 2022 B, Resp - BibliogYunior Sosa100% (1)
- Cardiología CTODocument95 paginiCardiología CTOEfren Penaflores100% (1)
- RMD 01 1819 Desgloses EdDocument9 paginiRMD 01 1819 Desgloses EdClaudia Familia100% (1)
- 03 Bioquimica DesglosesDocument4 pagini03 Bioquimica DesglosesElizabeth Cuevas Del RosarioÎncă nu există evaluări
- Desglose Cirugia AMIRDocument33 paginiDesglose Cirugia AMIRSiddhartha PalaciosÎncă nu există evaluări
- 04 - Cardiologia Desgloses Cto DominicanoDocument12 pagini04 - Cardiologia Desgloses Cto DominicanoMonica Rosario100% (2)
- CTO - Fisiología PDFDocument33 paginiCTO - Fisiología PDFcatoco12100% (3)
- CTO - HistologíaDocument33 paginiCTO - HistologíaJoha_Ade100% (2)
- Celina Curso EnurmDocument5 paginiCelina Curso EnurmPablo Francisco Montero TerreroÎncă nu există evaluări
- RMD 01 1819 Desgloses NFDocument4 paginiRMD 01 1819 Desgloses NFClaudia FamiliaÎncă nu există evaluări
- Desglose Enfermedades InfecciosasDocument18 paginiDesglose Enfermedades InfecciosasEstarlin Sosa JimenezÎncă nu există evaluări
- Respuestas Al Examen Mir 2015Document6 paginiRespuestas Al Examen Mir 2015wixon perezÎncă nu există evaluări
- Simulacro CtoDocument11 paginiSimulacro CtoRosse Julia Magno FigueredoÎncă nu există evaluări
- Desgloses NF PDFDocument14 paginiDesgloses NF PDFalma0pineda100% (1)
- Cirug (2Document239 paginiCirug (2Castro Brittany100% (1)
- Libro Gordo Ginecologia y ObstetriciaDocument39 paginiLibro Gordo Ginecologia y ObstetriciaGissela Yasmin Lozada FernandezÎncă nu există evaluări
- Banco Ciencias BásicasDocument40 paginiBanco Ciencias BásicasElviz LozadaÎncă nu există evaluări
- ADocument110 paginiAJames100% (1)
- Ginecología y ObstetriciaDocument12 paginiGinecología y ObstetriciaRAUL ORBEGOSO GAMBOAÎncă nu există evaluări
- UrologiaDocument11 paginiUrologiaMariela AlamoÎncă nu există evaluări
- Trabajo Sobre División Celular Biología César Márquez 3erob 31743432Document15 paginiTrabajo Sobre División Celular Biología César Márquez 3erob 31743432Adriana Ramírez CastilloÎncă nu există evaluări
- Guía Desarrollo Embrionariol1Document10 paginiGuía Desarrollo Embrionariol1Cinthia Lepe DiazÎncă nu există evaluări
- Monografía GametogénesisDocument23 paginiMonografía GametogénesisSaul Adrian Mamani SanchezÎncă nu există evaluări
- Manual CTO Ecuador 1º - EmbriologíaDocument32 paginiManual CTO Ecuador 1º - EmbriologíaAlexei100% (1)
- Monografía GametogénesisDocument23 paginiMonografía GametogénesisSaul Adrian Mamani SanchezÎncă nu există evaluări
- Guia Desarrollo Embrionario 2020Document10 paginiGuia Desarrollo Embrionario 2020Alejandro Rivera MenesesÎncă nu există evaluări
- EmbriologíaDocument9 paginiEmbriologíaNico RonquilloÎncă nu există evaluări
- Exp1 Desarrollo Del Sistema NerviosoDocument13 paginiExp1 Desarrollo Del Sistema Nerviosoalondra ÁvilaÎncă nu există evaluări
- James Diaz Cerón - Guia Pegagógica 8 Grado IERACV - Sistema Reproductor en HumanosDocument8 paginiJames Diaz Cerón - Guia Pegagógica 8 Grado IERACV - Sistema Reproductor en HumanosJames Norbayron Diaz CeronÎncă nu există evaluări
- HISTOLOGÍA - INFORME GRUPO 03 - Gametogénesis, Espermatogénesis y OvogénesisDocument17 paginiHISTOLOGÍA - INFORME GRUPO 03 - Gametogénesis, Espermatogénesis y OvogénesisMERLY DEILITH CUBAS GONZALESÎncă nu există evaluări
- Guía Desarrollo EmbrionarioDocument10 paginiGuía Desarrollo EmbrionarioHéctor Morales Ponce0% (1)
- Manual de Ontogenia Humana - EmbriologiaDocument43 paginiManual de Ontogenia Humana - EmbriologiaRicardo Yañez Adelaido100% (1)
- BL 13 Fecundación Desarrollo Embrionario y Fetal (V2)Document24 paginiBL 13 Fecundación Desarrollo Embrionario y Fetal (V2)jipk0% (1)
- Guia Morfo IIIDocument30 paginiGuia Morfo IIIvictoriaÎncă nu există evaluări
- RepasoembrionarioycortesDocument3 paginiRepasoembrionarioycortesMarta Corbo VillalbaÎncă nu există evaluări
- Desarrollo Embrionario Del Sistema NerviosoDocument17 paginiDesarrollo Embrionario Del Sistema NerviosoMabel PatiñoÎncă nu există evaluări
- Lara Alvarado SamaraDocument13 paginiLara Alvarado SamaraSam LaraÎncă nu există evaluări
- Taller Del Ciclo Menstrual y Las Etapas de Formación de Un Bebé TALLER SIENCIAS NATURALEZ...Document11 paginiTaller Del Ciclo Menstrual y Las Etapas de Formación de Un Bebé TALLER SIENCIAS NATURALEZ...deisyÎncă nu există evaluări
- Desarrollo EmbrionalDocument44 paginiDesarrollo EmbrionalPati González monteroÎncă nu există evaluări
- Biologia Humana - Actividad 2 MaquetaDocument10 paginiBiologia Humana - Actividad 2 Maquetamariana caceresÎncă nu există evaluări
- Examen de Pediatria Parte ADocument9 paginiExamen de Pediatria Parte APablo ParicahuaÎncă nu există evaluări
- Oferta Plazas Rem 2019 1Document193 paginiOferta Plazas Rem 2019 1Pablo ParicahuaÎncă nu există evaluări
- Preguntas Endocrinologia - Cuarta RotacionDocument4 paginiPreguntas Endocrinologia - Cuarta RotacionPablo ParicahuaÎncă nu există evaluări
- T6 IC Abdomen AgudoDocument17 paginiT6 IC Abdomen AgudoPablo ParicahuaÎncă nu există evaluări
- Herbario Sur PeruanoDocument1 paginăHerbario Sur PeruanoPablo ParicahuaÎncă nu există evaluări
- PDFDocument10 paginiPDFPablo Paricahua29% (7)
- DermaDocument6 paginiDermaPablo ParicahuaÎncă nu există evaluări
- Gestacionmultiple PDFDocument22 paginiGestacionmultiple PDFJair Eduardo De Jesus Crisanto BarretoÎncă nu există evaluări
- Capítulo 3 Primera Semana de Desarrollo de La Ovulación A La ImplantaciónDocument26 paginiCapítulo 3 Primera Semana de Desarrollo de La Ovulación A La ImplantaciónAndy GonzalezÎncă nu există evaluări
- Captura de Pantalla 2023-03-21 A La(s) 20.44.10Document102 paginiCaptura de Pantalla 2023-03-21 A La(s) 20.44.10Mariana SilvaÎncă nu există evaluări
- Amenorrea Primaria y SecundariaDocument31 paginiAmenorrea Primaria y SecundariaAlexis Delgado100% (1)
- Catálogo Modelos AnatómicosDocument22 paginiCatálogo Modelos AnatómicosRomulo FranciscoÎncă nu există evaluări
- Generalidades y Gametogénesis EstudianteDocument38 paginiGeneralidades y Gametogénesis Estudiantejuan andres arizmendiÎncă nu există evaluări
- Preguntas Tema 2 Estudio de La GestacionDocument2 paginiPreguntas Tema 2 Estudio de La GestacionTiffanyta1401Încă nu există evaluări
- EspermatogenesisDocument7 paginiEspermatogenesismariannyr13.02Încă nu există evaluări
- Prostata FemeninaDocument16 paginiProstata Femeninabigollo100% (1)
- Tesis-Erick-Correcciones FinalizadoDocument69 paginiTesis-Erick-Correcciones FinalizadoErick AlvarezÎncă nu există evaluări
- Exposicion Prolactina y LHDocument15 paginiExposicion Prolactina y LHKelly ArtÎncă nu există evaluări
- Anatomia Del Aparato Reproductor FemeninoDocument51 paginiAnatomia Del Aparato Reproductor FemeninoJen UculmanhaÎncă nu există evaluări
- Anatomía Sexual HumanaclsDocument14 paginiAnatomía Sexual HumanaclsMiguel Angel Berrocal VilcasÎncă nu există evaluări
- Anatomía Sexual HumanaDocument62 paginiAnatomía Sexual HumanaSebas PitanaÎncă nu există evaluări
- Aparato Reproductor FemeninoDocument26 paginiAparato Reproductor FemeninoAssiral MendozaÎncă nu există evaluări
- ACTIVIDAD Aparato Reproductor Masculino y FemeninoDocument3 paginiACTIVIDAD Aparato Reproductor Masculino y Femeninodalila rueda harrisÎncă nu există evaluări
- MalformacionDocument13 paginiMalformacionValeria VillalobosÎncă nu există evaluări
- Balotario de PreguntasDocument6 paginiBalotario de PreguntasGustavo Alexander Zapana QuispeÎncă nu există evaluări
- Preguntas Gineco 1ra Clas NG PDFDocument1 paginăPreguntas Gineco 1ra Clas NG PDFmela1997Încă nu există evaluări
- NEUROENDOCRINODocument12 paginiNEUROENDOCRINOYARIEL ALONSO ALVAREZÎncă nu există evaluări
- Embriologia y Anatomia Aparato Reproductor Femenino y Mama UpaoDocument65 paginiEmbriologia y Anatomia Aparato Reproductor Femenino y Mama Upaofernando cruz diosesÎncă nu există evaluări
- Eje Hipotálamo Hipófisis GonadalDocument3 paginiEje Hipotálamo Hipófisis GonadalPao Pao Z100% (1)
- Desarrollo Normal Del AdolescenteDocument7 paginiDesarrollo Normal Del AdolescenteAlejandro EspinozaÎncă nu există evaluări
- CIENCIAS NATURALES. Aparato Reproductor Femenino y MasculinoDocument9 paginiCIENCIAS NATURALES. Aparato Reproductor Femenino y MasculinoGi GüizzoÎncă nu există evaluări
- Evaluacion Pubertad Sexto BasicoDocument6 paginiEvaluacion Pubertad Sexto BasicoJessica SilvaÎncă nu există evaluări
- PROYECTO INTERDISCIPLINAR 7nDocument16 paginiPROYECTO INTERDISCIPLINAR 7nTC TomalasitaÎncă nu există evaluări
- Esperma To Bioscop I ADocument1 paginăEsperma To Bioscop I AJorge E. Ramirez NarvaezÎncă nu există evaluări
- Cuadro Comparativo Aparato ReproductorDocument3 paginiCuadro Comparativo Aparato ReproductorYessica AguilarÎncă nu există evaluări
- Aparato GenitalDocument51 paginiAparato GenitalJOSELIN MILAGROS RAMOS HUANCAÎncă nu există evaluări
- Ciclo MestrualDocument16 paginiCiclo MestrualPatty Vasquez100% (3)
- Suprelorin Dosier TecnicoDocument33 paginiSuprelorin Dosier TecnicoJudithÎncă nu există evaluări