S-ar putea să vă placă și
- Beethoven Piano Sonata Op. 57 N. 23Document29 paginiBeethoven Piano Sonata Op. 57 N. 23GiancarloÎncă nu există evaluări
- Post Purchase Cognitive DissonanceDocument18 paginiPost Purchase Cognitive DissonancePawan DihiyeÎncă nu există evaluări
- Proposed USG Boral Ceiling System For Tropicana MiyuDocument5 paginiProposed USG Boral Ceiling System For Tropicana MiyuLorraineÎncă nu există evaluări
- GP150 PDFDocument125 paginiGP150 PDFBf Ipanema100% (1)
- Constitutions and ConstitutionalismDocument10 paginiConstitutions and ConstitutionalismmylovebhuÎncă nu există evaluări
- A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in PlantsDocument3 paginiA Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plantstanuarora09Încă nu există evaluări
- 1 s2.0 S0735109715029083 MainDocument2 pagini1 s2.0 S0735109715029083 Mainwelly yantoÎncă nu există evaluări
- Forsberg Et Al 1994 Use of Transcriptional Fusions To Monitor Gene Expression A Cautionary TaleDocument5 paginiForsberg Et Al 1994 Use of Transcriptional Fusions To Monitor Gene Expression A Cautionary TaleKunal KumarÎncă nu există evaluări
- Clase 22 Febrero 2016 Transcripcion TraduccionDocument52 paginiClase 22 Febrero 2016 Transcripcion TraduccionAlma A OrtizÎncă nu există evaluări
- The Role of Microrna-15A in The Development of Prostate Cancer - Effects On Cell Proliferation and Pro-Inflammatory SignallingDocument5 paginiThe Role of Microrna-15A in The Development of Prostate Cancer - Effects On Cell Proliferation and Pro-Inflammatory SignallingTeodorÎncă nu există evaluări
- 2686 PDFDocument11 pagini2686 PDFmojÎncă nu există evaluări
- Intracellular Hepatitis C Virus RNA-dependent RNA Polymerase ActivityDocument4 paginiIntracellular Hepatitis C Virus RNA-dependent RNA Polymerase ActivitybmnlthyukÎncă nu există evaluări
- Dex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisDocument6 paginiDex3 and Survivin-2B: Two Novel Splice Variants of The ApoptosisNona NonicaaÎncă nu există evaluări
- 2309 Full PDFDocument10 pagini2309 Full PDFagus karsetiyonoÎncă nu există evaluări
- Inactivating Mutations of Caspase-8 Gene in Colorectal CarcinomasDocument8 paginiInactivating Mutations of Caspase-8 Gene in Colorectal Carcinomasopus.mj1Încă nu există evaluări
- PMA Up-Regulates The Transcription of Axl by AP-1 Transcription Factor Binding To TRE Sequences Via The MAPK Cascade in Leukaemia CellsDocument19 paginiPMA Up-Regulates The Transcription of Axl by AP-1 Transcription Factor Binding To TRE Sequences Via The MAPK Cascade in Leukaemia CellsWilliam RamirezÎncă nu există evaluări
- Zhong 1999Document4 paginiZhong 1999Araceli Enríquez OvandoÎncă nu există evaluări
- Ijcep0007 0602Document9 paginiIjcep0007 0602Deedee RenovaldiÎncă nu există evaluări
- Artículo ViroDocument13 paginiArtículo ViroNatalia BogotáÎncă nu există evaluări
- Adapted Tembusu-Like Virus in Chickens and Geese in ChinaDocument3 paginiAdapted Tembusu-Like Virus in Chickens and Geese in ChinaTrung Kien LeÎncă nu există evaluări
- Science AbmDocument7 paginiScience AbmCosti ValiÎncă nu există evaluări
- Knockdown Genico Especifico y Eficiente Por ARNsi Producidos Por Bacterias Huang Et Al 2013Document9 paginiKnockdown Genico Especifico y Eficiente Por ARNsi Producidos Por Bacterias Huang Et Al 2013valenciabastoÎncă nu există evaluări
- Somatic Protein ArticleDocument9 paginiSomatic Protein ArticleVanessa de AndradeÎncă nu există evaluări
- 4030 FullDocument10 pagini4030 FullKartikeya SinghÎncă nu există evaluări
- Biology of CancerDocument62 paginiBiology of CancerSavitaÎncă nu există evaluări
- SignallingDocument74 paginiSignallingAshrafÎncă nu există evaluări
- Revised EditionDocument19 paginiRevised Editionfafy mohammedÎncă nu există evaluări
- Chromosomal Aberrations Induced by 5-Azacytidine Combined With VP-16 (Etoposide) in CHO-K1 and XRS-5 Cell LinesDocument16 paginiChromosomal Aberrations Induced by 5-Azacytidine Combined With VP-16 (Etoposide) in CHO-K1 and XRS-5 Cell LinesAry AguiarÎncă nu există evaluări
- Autoantibodies in P45011D6, Cytochrome: LKM-1 Recognize Short Sequence P-450 MonooxygenaseDocument9 paginiAutoantibodies in P45011D6, Cytochrome: LKM-1 Recognize Short Sequence P-450 MonooxygenasefryzinÎncă nu există evaluări
- Reverse Transcription of The Ribonucleic AcidDocument2 paginiReverse Transcription of The Ribonucleic AcidOmeyya TanveerÎncă nu există evaluări
- The Genetic Background of Pancreatic Cancer - Genes That Might Be Biomarkers or Indicators of Metastasis To The Lung.Document1 paginăThe Genetic Background of Pancreatic Cancer - Genes That Might Be Biomarkers or Indicators of Metastasis To The Lung.Laur NeyÎncă nu există evaluări
- Kenck 1996Document5 paginiKenck 1996Sarly FebrianaÎncă nu există evaluări
- Cancer: When Good Cells Go BadDocument62 paginiCancer: When Good Cells Go BadEpi Panjaitan100% (1)
- 1 s2.0 S0014482702955813 MainDocument12 pagini1 s2.0 S0014482702955813 MainKevin MaiseyÎncă nu există evaluări
- Thomas 2007Document11 paginiThomas 2007edelinÎncă nu există evaluări
- In - Vivo 33 777Document9 paginiIn - Vivo 33 777ingelitaÎncă nu există evaluări
- The Transcription Factor ZBP-89 Controls Generation of The Hematopoietic Lineage in Zebrafish and Mouse Embryonic Stem CellsDocument10 paginiThe Transcription Factor ZBP-89 Controls Generation of The Hematopoietic Lineage in Zebrafish and Mouse Embryonic Stem CellsNeelra JeanloÎncă nu există evaluări
- Loss of P16 in Esophageal Adenocarcinoma Detected by Fluorescence in Situ Hybridization and ImmunohistochemistryDocument6 paginiLoss of P16 in Esophageal Adenocarcinoma Detected by Fluorescence in Situ Hybridization and ImmunohistochemistryTeodorÎncă nu există evaluări
- Int. Immunol.-1999-Staal-317-23Document7 paginiInt. Immunol.-1999-Staal-317-23Karthik AripakaÎncă nu există evaluări
- Abnormal Spermatogenesis and Reduced Fertility in Transition Nuclear Protein 1-Deficient MiceDocument6 paginiAbnormal Spermatogenesis and Reduced Fertility in Transition Nuclear Protein 1-Deficient MiceBerlinÎncă nu există evaluări
- Predictive Value of Mirna 181a in PediatricDocument1 paginăPredictive Value of Mirna 181a in Pediatriclaura rangelÎncă nu există evaluări
- Rickettsias: Angelica Barbosa BuitragoDocument63 paginiRickettsias: Angelica Barbosa BuitragoLeo LopezÎncă nu există evaluări
- Differential Display RT PCR of Total RNA From Human Foreskin Fibroblasts For Investigation of Androgen-Dependent Gene ExpressionDocument8 paginiDifferential Display RT PCR of Total RNA From Human Foreskin Fibroblasts For Investigation of Androgen-Dependent Gene ExpressionAltaicaÎncă nu există evaluări
- PlasmidsDocument12 paginiPlasmidsMaciel RodriguezÎncă nu există evaluări
- Cells Preferentially Vp3 And/or: T in Psoriatic Lesions Use T-Cell Receptor GenesDocument15 paginiCells Preferentially Vp3 And/or: T in Psoriatic Lesions Use T-Cell Receptor GenesSnehal BhagatÎncă nu există evaluări
- The Expression of Annexin II and Its Role in The Fibrinolytic Activity in Acute Promyelocytic LeukemiaDocument6 paginiThe Expression of Annexin II and Its Role in The Fibrinolytic Activity in Acute Promyelocytic LeukemiaLaura LópezÎncă nu există evaluări
- Research Epstein-Barr Virus Encoded Latent MembraneDocument9 paginiResearch Epstein-Barr Virus Encoded Latent MembraneMiss AmyÎncă nu există evaluări
- CoverDocument7 paginiCoveryasmineÎncă nu există evaluări
- Biochimie Volume 81 Issue 6-Supp-S1 1999 (Doi 10.1016/s0300-9084 (99) 80112-6) - The Sir Hans Krebs LectureDocument390 paginiBiochimie Volume 81 Issue 6-Supp-S1 1999 (Doi 10.1016/s0300-9084 (99) 80112-6) - The Sir Hans Krebs LectureAllcrissÎncă nu există evaluări
- Flejter 1996Document6 paginiFlejter 1996Rahul AmbawataÎncă nu există evaluări
- Janssen 2010Document4 paginiJanssen 2010barti koksÎncă nu există evaluări
- ASC Abstracts 05Document110 paginiASC Abstracts 0512345qwertasdfgzxcvbÎncă nu există evaluări
- Expression of iNOS in Oral Mucosal Cells by RT-PCR AND ImmunocytochemistryDocument6 paginiExpression of iNOS in Oral Mucosal Cells by RT-PCR AND ImmunocytochemistryrizwanÎncă nu există evaluări
- Leucemia Promielocitica AgudaDocument11 paginiLeucemia Promielocitica AgudaJulio AtahualpaÎncă nu există evaluări
- Embo Embo Embo: C-Secretase ActivityDocument11 paginiEmbo Embo Embo: C-Secretase ActivityMaria Vonny WijayaÎncă nu există evaluări
- Señalización Molecular: Julio Hilario VargasDocument43 paginiSeñalización Molecular: Julio Hilario VargasAndrew Emilio Castillo PachecoÎncă nu există evaluări
- Duncan 2000Document4 paginiDuncan 2000Silvia LascarezÎncă nu există evaluări
- The Pseudomonas Chlororaphis PCL1391 Sigma Regulator Psra Represses The Production of The Antifungal Metabolite Phenazine-1-Carboxamide PDFDocument10 paginiThe Pseudomonas Chlororaphis PCL1391 Sigma Regulator Psra Represses The Production of The Antifungal Metabolite Phenazine-1-Carboxamide PDFNabil Mahesa FahmiÎncă nu există evaluări
- Fujioka 1999Document9 paginiFujioka 1999Sharon ChiangÎncă nu există evaluări
- Oncogene: Viral OncogenesDocument12 paginiOncogene: Viral OncogenesIlyas Khan AurakzaiÎncă nu există evaluări
- Tian 2016Document13 paginiTian 2016Epiphany SonderÎncă nu există evaluări
- Systemic Biodistribution Expression Mice: Gene Therapy: and Long-Term of A Transgene inDocument5 paginiSystemic Biodistribution Expression Mice: Gene Therapy: and Long-Term of A Transgene inCa Nd RaÎncă nu există evaluări
- Edwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Document48 paginiEdwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Edwin DarmawanÎncă nu există evaluări
- ARTICULO Transposable Elem CancerDocument5 paginiARTICULO Transposable Elem CancerJuanfer Bonett OntanedaÎncă nu există evaluări
- Terapia Sistémica en Carcinoma Metastásico de Colon Y Recto: Mauricio Lema Medina MDDocument14 paginiTerapia Sistémica en Carcinoma Metastásico de Colon Y Recto: Mauricio Lema Medina MDapi-26302710Încă nu există evaluări
- Nature 07385Document8 paginiNature 07385api-26302710Încă nu există evaluări
- For Refractory Cutaneous T-Cell Lymphoma (CTCL) Phase 2 Trial of Oral Vorinostat (Suberoylanilide Hydroxamic Acid, SAHA)Document10 paginiFor Refractory Cutaneous T-Cell Lymphoma (CTCL) Phase 2 Trial of Oral Vorinostat (Suberoylanilide Hydroxamic Acid, SAHA)api-26302710Încă nu există evaluări
- Mycosis Fungoides and Sézary Syndrome: SeminarDocument13 paginiMycosis Fungoides and Sézary Syndrome: Seminarapi-26302710Încă nu există evaluări
- Guias para Administrar en Forma Segura Altas Dosis de Interleukina 2007Document2 paginiGuias para Administrar en Forma Segura Altas Dosis de Interleukina 2007api-26302710Încă nu există evaluări
- Aldesleukin For Injection RX Only: DescriptionDocument16 paginiAldesleukin For Injection RX Only: Descriptionapi-26302710Încă nu există evaluări
- Ncologist: R I FDADocument7 paginiNcologist: R I FDAapi-26302710Încă nu există evaluări
- Ca Screening 2008Document20 paginiCa Screening 2008api-26302710Încă nu există evaluări
- CaScreening2009 01Document16 paginiCaScreening2009 01api-26302710Încă nu există evaluări
- Molecular Targeted Therapy of Cancer: Brain TumorsDocument33 paginiMolecular Targeted Therapy of Cancer: Brain Tumorsapi-26302710Încă nu există evaluări
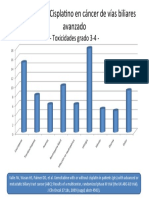
- Gem Cis ABC02Document1 paginăGem Cis ABC02api-26302710Încă nu există evaluări
- Sutent GistDocument10 paginiSutent Gistapi-26302710Încă nu există evaluări
- Controversiesinthemanagementof Relapsedandprimaryrefractory Hodgkin'SlymphomaDocument8 paginiControversiesinthemanagementof Relapsedandprimaryrefractory Hodgkin'Slymphomaapi-26302710Încă nu există evaluări
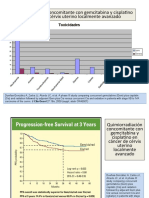
- Gem Cis RTin Ca CervixDocument3 paginiGem Cis RTin Ca Cervixapi-26302710Încă nu există evaluări
- Lysosomal Storage Disease 2: SeriesDocument12 paginiLysosomal Storage Disease 2: Seriesapi-26302710Încă nu există evaluări
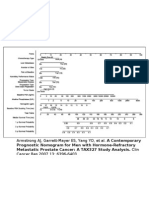
- HRPC Nomogram97Document1 paginăHRPC Nomogram97api-26302710Încă nu există evaluări
- Acer AL1516 Service Guide: Downloaded From Manuals Search EngineDocument48 paginiAcer AL1516 Service Guide: Downloaded From Manuals Search Enginerudra_1Încă nu există evaluări
- The BoarderDocument3 paginiThe BoarderAnonymous iX6KV9LZzoÎncă nu există evaluări
- Exercises Problem 2 Lesson 4Document2 paginiExercises Problem 2 Lesson 4Marianne DevillenaÎncă nu există evaluări
- Nakul VermaDocument112 paginiNakul VermaNakul VermaÎncă nu există evaluări
- Application Note Assessing Flow Accelerated Corrosion With Pulsed Eddy CurrentDocument2 paginiApplication Note Assessing Flow Accelerated Corrosion With Pulsed Eddy CurrentZoran TripunovskiÎncă nu există evaluări
- S-Block Elements: Earth Metals. These Are So Called Because Their Oxides and Hydroxides Are Alkaline in NatureDocument8 paginiS-Block Elements: Earth Metals. These Are So Called Because Their Oxides and Hydroxides Are Alkaline in NatureAgamGoelÎncă nu există evaluări
- Computer Commerce PDFDocument176 paginiComputer Commerce PDFMallu TechÎncă nu există evaluări
- Quiz ErrorsDocument5 paginiQuiz Errorsanon_843459121Încă nu există evaluări
- Mock MU-BAAT Final Version Set 1Document8 paginiMock MU-BAAT Final Version Set 1Priyanshu SinghÎncă nu există evaluări
- Father William - Poem by Lewis CarrollDocument1 paginăFather William - Poem by Lewis CarrollRachid AfrznaneÎncă nu există evaluări
- Summative Test Module 1.1 1.2Document2 paginiSummative Test Module 1.1 1.2Jocelyn RoxasÎncă nu există evaluări
- Substantive Testing in The Revenue CycleDocument3 paginiSubstantive Testing in The Revenue CycleGeorgia FlorentinoÎncă nu există evaluări
- WordDocument48 paginiWordenkelene.zulfajÎncă nu există evaluări
- Brand Identity Aaker's ModelDocument2 paginiBrand Identity Aaker's Modelvibhaanagpal100% (1)
- Astm A490M Heavy Hex Structural Bolts (Metric)Document7 paginiAstm A490M Heavy Hex Structural Bolts (Metric)cragmarÎncă nu există evaluări
- Christocentric Theology: A Theology Shaped by Faith in The Person, Teachings, and Work of Jesus ChristDocument9 paginiChristocentric Theology: A Theology Shaped by Faith in The Person, Teachings, and Work of Jesus ChristDrew Dixon100% (1)
- Fieldwork Exercise 2: 4 Thailand Geography Olympiad Chiang Mai University Tuesday, 10 May 2022 Time: 13:30-16:30Document12 paginiFieldwork Exercise 2: 4 Thailand Geography Olympiad Chiang Mai University Tuesday, 10 May 2022 Time: 13:30-16:30Sun CharoenratÎncă nu există evaluări
- Manual Stiga VILLA 92 MDocument6 paginiManual Stiga VILLA 92 MCzesław WierzchowskiÎncă nu există evaluări
- Johnson Power Pumping CatalogDocument24 paginiJohnson Power Pumping CatalogAlvaro Patricio Etcheverry TroncosoÎncă nu există evaluări
- Lembar Observasi Kimia PKL 2023Document41 paginiLembar Observasi Kimia PKL 2023Nur Sri WahyuniÎncă nu există evaluări
- Surf Excel ND AirelDocument13 paginiSurf Excel ND AirelPratibha Chandil100% (1)
- Unit 2 Computer Hardware and SoftwareDocument24 paginiUnit 2 Computer Hardware and SoftwareNgHanSeongÎncă nu există evaluări
- Escalation Estimating Principles and Methods Using IndicesDocument4 paginiEscalation Estimating Principles and Methods Using IndicesAlejandro RuizÎncă nu există evaluări
- Odisha PWD Registration in CDMSDocument2 paginiOdisha PWD Registration in CDMSSabyasachi Naik (Zico)Încă nu există evaluări
- Design and Implementation of A Turbo Code System On FPGA: November 2011Document6 paginiDesign and Implementation of A Turbo Code System On FPGA: November 2011Ankit GuptaÎncă nu există evaluări
- Tryout Un Bahasa Inggris SMKDocument9 paginiTryout Un Bahasa Inggris SMKSMK Miftahul HikmahÎncă nu există evaluări