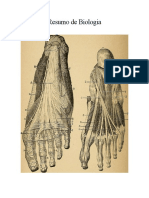
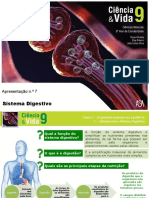
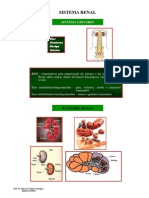
S-ar putea să vă placă și
- 3º Ano Atividades Da 15 Semana Port.e CienciasDocument11 pagini3º Ano Atividades Da 15 Semana Port.e CienciasKARLA FERREIRA DOS SANTOS100% (14)
- Epidemiologia Manual Versao 2014aDocument35 paginiEpidemiologia Manual Versao 2014aMarcelo Cezar SoaresÎncă nu există evaluări
- Manual de Boas PraticasDocument44 paginiManual de Boas PraticasEdy100% (3)
- Manual de Inspesao e Controle de Qualidade Do LeiteDocument101 paginiManual de Inspesao e Controle de Qualidade Do LeiteJuliana Alexandre100% (1)
- Fisiologia Dos Cordados - Questões Com GabaritoDocument25 paginiFisiologia Dos Cordados - Questões Com GabaritoPaulo HenriqueÎncă nu există evaluări
- A BiosferaDocument85 paginiA BiosferaMiriam CarvalheiroÎncă nu există evaluări
- Classificação Dos Seres Vivos TextoDocument6 paginiClassificação Dos Seres Vivos TextoPatrícia Alexandra da Silva Paz100% (1)
- Cadeias e Teias AlimentaresDocument3 paginiCadeias e Teias AlimentaresJoceli Maria ZandonaiÎncă nu există evaluări
- Fermentação e RespiraçãoDocument20 paginiFermentação e RespiraçãoRafaela Afonso100% (1)
- Dissertativas UEGDocument36 paginiDissertativas UEGLucileyÎncă nu există evaluări
- Classificação Dos Seres Vivos PDFDocument3 paginiClassificação Dos Seres Vivos PDFGiseleDuvalÎncă nu există evaluări
- c6 Curso B Prof BiologiaDocument28 paginic6 Curso B Prof BiologiaHancock86Încă nu există evaluări
- Chondrichthyes GeralDocument104 paginiChondrichthyes GeralCelso Cardeal Jr.67% (3)
- 152 Sistema Digestorio - Resumo PDFDocument2 pagini152 Sistema Digestorio - Resumo PDFLandroÎncă nu există evaluări
- Atividade Da ProfessoraDocument4 paginiAtividade Da ProfessoraBrendo Calado da SilvaÎncă nu există evaluări
- O Sistema DigestivoDocument5 paginiO Sistema DigestivoBruna Alexandra MartinsÎncă nu există evaluări
- SISTEMA MuscularDocument2 paginiSISTEMA MuscularPatricia HerbsterÎncă nu există evaluări
- Origem e Evolucao Do Plano Corpóreo de Gastropodes 01Document11 paginiOrigem e Evolucao Do Plano Corpóreo de Gastropodes 01Vanessa Bezerra100% (3)
- Trajano E. Osteichthyes, Actinopterygii PDFDocument29 paginiTrajano E. Osteichthyes, Actinopterygii PDFFábio André Facco JacomassaÎncă nu există evaluări
- MMMMMMMDocument6 paginiMMMMMMMMaisaOliveiraÎncă nu există evaluări
- Teste 6Document3 paginiTeste 6Allan Gimenes100% (1)
- 40 Obtenção de Energia - FermentaçãoDocument43 pagini40 Obtenção de Energia - FermentaçãoAna Filipa Fernandes FigueiredoÎncă nu există evaluări
- Características Dos Seres VivosDocument46 paginiCaracterísticas Dos Seres VivosOtilia Marcelino AfonsoÎncă nu există evaluări
- Citoplasma e NúcleoDocument11 paginiCitoplasma e NúcleoAna Paula Porto FerreiraÎncă nu există evaluări
- BIO Fuvest PDFDocument115 paginiBIO Fuvest PDFMaria Eduarda Moraes IampietroÎncă nu există evaluări
- Diversidade Na BiosferaDocument73 paginiDiversidade Na BiosferaDino RubioÎncă nu există evaluări
- Respiração Aeróbia Anaeróbia e FotossínteseDocument7 paginiRespiração Aeróbia Anaeróbia e FotossínteseRanielle Montanini100% (3)
- Biologia - Pré-Vestibular Vetor - Bio1 Fisiologia - Sistema EndócrinoDocument3 paginiBiologia - Pré-Vestibular Vetor - Bio1 Fisiologia - Sistema EndócrinoBiologia Concurso Vestibular100% (5)
- Matéria e Energia No EcossistemaDocument6 paginiMatéria e Energia No EcossistemaFelipe AlcantaraÎncă nu există evaluări
- Apostila de EcologiaDocument15 paginiApostila de EcologiaJoão PedroÎncă nu există evaluări
- Polonio PDFDocument2 paginiPolonio PDFwozikÎncă nu există evaluări
- Lista de Exercício para Revisão AT5 - 2 SÉRIEDocument7 paginiLista de Exercício para Revisão AT5 - 2 SÉRIEPedro Dorea ff almeidaÎncă nu există evaluări
- GLICOCONJUGADOSDocument1 paginăGLICOCONJUGADOSLiquinho CanutoÎncă nu există evaluări
- Origem e Evolução Das Células - Texto 1 - Metodologia AtivaDocument2 paginiOrigem e Evolução Das Células - Texto 1 - Metodologia Ativaabraaobiologo@gmail.comÎncă nu există evaluări
- Aula PolialeliaDocument14 paginiAula PolialeliaSavio Sa100% (1)
- Ficha Informativa Fluxo de Energia e Ciclo Da Matéria 8ºano - 22 - 23Document5 paginiFicha Informativa Fluxo de Energia e Ciclo Da Matéria 8ºano - 22 - 23Pedro Costa(520)Încă nu există evaluări
- Exercicio 1 GenéticaDocument3 paginiExercicio 1 GenéticaThássio MouraÎncă nu există evaluări
- Filogenia MolecularDocument25 paginiFilogenia MolecularkirleymichellyÎncă nu există evaluări
- Os Grandes Filos Dos MetazoáriosDocument42 paginiOs Grandes Filos Dos MetazoáriosEmerson MendesÎncă nu există evaluări
- Atividade CladogramaDocument6 paginiAtividade CladogramaArthurÎncă nu există evaluări
- 4.1.1. Ingestão, Digestão e Absorção em Seres Vivos HeterotróficosDocument20 pagini4.1.1. Ingestão, Digestão e Absorção em Seres Vivos HeterotróficosClashGamerÎncă nu există evaluări
- Cad C3 3ano 4opcao Tarefa BiologiaDocument16 paginiCad C3 3ano 4opcao Tarefa BiologiaFyby almeidaÎncă nu există evaluări
- Exercicios Fluxo EnergéticoDocument2 paginiExercicios Fluxo EnergéticoLucas OrtolanÎncă nu există evaluări
- Texto Ecologia 2º AnoDocument6 paginiTexto Ecologia 2º AnoEllenÎncă nu există evaluări
- Panarthropoda - Introdução Aos ArthropodaDocument9 paginiPanarthropoda - Introdução Aos ArthropodaLourdes Elmoor LoureiroÎncă nu există evaluări
- Embriologia e HistologiaDocument5 paginiEmbriologia e HistologiaBruna YukaÎncă nu există evaluări
- Exercício Sobre EnergiaDocument6 paginiExercício Sobre EnergiaAndrey SiqueiraÎncă nu există evaluări
- Introdução À Medicina Laboratorial MoçambiqueDocument61 paginiIntrodução À Medicina Laboratorial MoçambiqueFolario Draiva Tefo100% (1)
- Atividade 68742Document5 paginiAtividade 68742Gelma XavierÎncă nu există evaluări
- Resumo - Zoologia de InvertebradosDocument70 paginiResumo - Zoologia de InvertebradosJocilene RodriguesÎncă nu există evaluări
- Resumo de BiologiaDocument6 paginiResumo de BiologiaFranciscoÎncă nu există evaluări
- O Reino AnimaliaDocument5 paginiO Reino AnimaliaInfospace100% (1)
- Aula 1 Origem Dos CloroplastosDocument54 paginiAula 1 Origem Dos CloroplastosTania RuizÎncă nu există evaluări
- Ciclos BiogeoquímicosDocument18 paginiCiclos BiogeoquímicosRafaela MartinsÎncă nu există evaluări
- Atividades de Sistema DigestorioDocument5 paginiAtividades de Sistema DigestorioJackelineNunesÎncă nu există evaluări
- C3 CursoE Biologia B 20aulasDocument12 paginiC3 CursoE Biologia B 20aulasHancock86Încă nu există evaluări
- Origem e Radiação Das Angiosperm As - Alessandro Rapini 2003Document60 paginiOrigem e Radiação Das Angiosperm As - Alessandro Rapini 2003Floyder100% (2)
- Fluxo de Matéria e EnergiaDocument17 paginiFluxo de Matéria e EnergiaAna Flávia100% (1)
- Especiação e FilogeniaDocument3 paginiEspeciação e FilogeniaVanessa NogueiraÎncă nu există evaluări
- Sistema DigestivoDocument7 paginiSistema DigestivoAna Catarina Yehorova Rodrigues Quiterio (9 1 SV)Încă nu există evaluări
- Mecanismos de EspeciaçãoDocument8 paginiMecanismos de EspeciaçãoCamila de QuadrosÎncă nu există evaluări
- Função e Disfunção 3 - Digestão Aves - EquinosDocument9 paginiFunção e Disfunção 3 - Digestão Aves - EquinosSítio das CalopsitasÎncă nu există evaluări
- Digestão Geral Dos AnimaisDocument17 paginiDigestão Geral Dos AnimaisThiago PortesÎncă nu există evaluări
- Sist. Digest. Das AvesDocument4 paginiSist. Digest. Das Avessammyprado13Încă nu există evaluări
- Banco de Imagens EmbrapaDocument1 paginăBanco de Imagens EmbrapaJuliana AlexandreÎncă nu există evaluări
- Livro - Anatomia e Fisiologia Dos Peixes de Agua DoceDocument49 paginiLivro - Anatomia e Fisiologia Dos Peixes de Agua DoceAkio MarcolaÎncă nu există evaluări
- Açaí Microbiologia 3Document5 paginiAçaí Microbiologia 3Juliana AlexandreÎncă nu există evaluări
- Código Sanitário TimonDocument101 paginiCódigo Sanitário TimonJuliana AlexandreÎncă nu există evaluări
- InfelizDocument17 paginiInfelizJuliana AlexandreÎncă nu există evaluări
- Açaí AntioxidanteDocument7 paginiAçaí AntioxidanteJuliana AlexandreÎncă nu există evaluări
- 9 21 1 PBDocument13 pagini9 21 1 PBEduardo SilvaÎncă nu există evaluări
- Limpeza de VidrariasDocument6 paginiLimpeza de VidrariasGrace FernandesÎncă nu există evaluări
- Artigo Vigilancia Sanitária de Alimentos - RevisãoDocument25 paginiArtigo Vigilancia Sanitária de Alimentos - RevisãoMikaelle PedrosaÎncă nu există evaluări
- Suínos Teoria e Prática PDFDocument908 paginiSuínos Teoria e Prática PDFLucilene Martins67% (3)
- Biologia - Apostila ANVISA - Módulo 04Document66 paginiBiologia - Apostila ANVISA - Módulo 04Biologia ANVISA100% (3)
- F 02 PL BTDocument761 paginiF 02 PL BTJuliana AlexandreÎncă nu există evaluări
- Qualidade Industria de Alimentos PDFDocument46 paginiQualidade Industria de Alimentos PDFGaldirJuniorÎncă nu există evaluări
- Pag26 32Document7 paginiPag26 32Juliana AlexandreÎncă nu există evaluări
- 7 - Elaine Alves Dos SantosDocument162 pagini7 - Elaine Alves Dos SantosJuliana AlexandreÎncă nu există evaluări
- Sujidades AçaiDocument2 paginiSujidades AçaiJuliana AlexandreÎncă nu există evaluări
- In 62 2011Document24 paginiIn 62 2011Jana Kelly SantosÎncă nu există evaluări
- 20 Orientacoes para o Controle LeiteiroDocument2 pagini20 Orientacoes para o Controle LeiteiroJuliana AlexandreÎncă nu există evaluări
- AW Obstetrícia Apostila 2 PDFDocument38 paginiAW Obstetrícia Apostila 2 PDFJuliana AlexandreÎncă nu există evaluări
- 2009 FabioHenriqueBezerraXimenesDocument66 pagini2009 FabioHenriqueBezerraXimenesJuliana AlexandreÎncă nu există evaluări
- Infecções CirúrgicasDocument7 paginiInfecções CirúrgicasJuliana AlexandreÎncă nu există evaluări
- Aula Fos FoxDocument31 paginiAula Fos FoxJuliana AlexandreÎncă nu există evaluări
- 7 - RenalDocument9 pagini7 - RenalJuliana AlexandreÎncă nu există evaluări
- Diana Gabaldon - Outlander 06 - Um Sopro de Neve e Cinzas PDFDocument855 paginiDiana Gabaldon - Outlander 06 - Um Sopro de Neve e Cinzas PDFAnia Arievilo100% (2)
- Aula 02 - PelvimetriaDocument3 paginiAula 02 - PelvimetriaJuliana AlexandreÎncă nu există evaluări
- Células de FungosDocument2 paginiCélulas de FungosJuliana AlexandreÎncă nu există evaluări
- Quebra VentosDocument6 paginiQuebra Ventosveleda9734Încă nu există evaluări
- Guia Nacional de Benefícios 2021Document93 paginiGuia Nacional de Benefícios 2021Rh MetalicaÎncă nu există evaluări
- Erosão Do Solo Atividades 3º Ou 4º AnosDocument2 paginiErosão Do Solo Atividades 3º Ou 4º AnosValdeci Sousa FrançaÎncă nu există evaluări
- Fadiga VisualDocument6 paginiFadiga VisualvagnerÎncă nu există evaluări
- PsicologiaDocument31 paginiPsicologiaJaqueline Nascimento JS100% (2)
- Artigo 3 - Fundamentos Da Cosmetologia - Sabonetes de GlicerinaDocument8 paginiArtigo 3 - Fundamentos Da Cosmetologia - Sabonetes de GlicerinaAna CarolinaÎncă nu există evaluări
- Fratura de PelveDocument97 paginiFratura de PelveFlavio DucciniÎncă nu există evaluări
- Lista 2 - Ft2Document3 paginiLista 2 - Ft2Emily FreireÎncă nu există evaluări
- Manual de Fiscalizacao de Pescado e Derivados v1.2021Document49 paginiManual de Fiscalizacao de Pescado e Derivados v1.2021Amanda HervisÎncă nu există evaluări
- Dotações de Enfermagem Impacte Resultados em Saude PDFDocument118 paginiDotações de Enfermagem Impacte Resultados em Saude PDFTiagoBarretoÎncă nu există evaluări
- Caderno Do Despertar EmocionalDocument18 paginiCaderno Do Despertar EmocionalLuiz Carlos FotografoÎncă nu există evaluări
- Bactérias Gram - PositivasDocument12 paginiBactérias Gram - PositivasLúria RochaÎncă nu există evaluări
- 941x025-6-7-8 Isometsolar Solucoes v2Document4 pagini941x025-6-7-8 Isometsolar Solucoes v2joaotgilÎncă nu există evaluări
- Roteiro - Autismo PDFDocument4 paginiRoteiro - Autismo PDFHelaine BorgesÎncă nu există evaluări
- Folheto Cat14Document5 paginiFolheto Cat14joaoÎncă nu există evaluări
- Pensos ql2013Document24 paginiPensos ql2013Alexandre MedeirosÎncă nu există evaluări
- Relatório Sobre Câncer BucalDocument5 paginiRelatório Sobre Câncer BucalGEOVANNA KETLLY TELES DOS SANTOSÎncă nu există evaluări
- Simulado AV Geração de Energia Elétrica EstácioDocument4 paginiSimulado AV Geração de Energia Elétrica EstácioadrianoÎncă nu există evaluări
- 2008 Termo de Referencia - Relatorio de Controle Ambiental RcaDocument4 pagini2008 Termo de Referencia - Relatorio de Controle Ambiental RcaRaquel MenezesÎncă nu există evaluări
- Resolucao 828 2020 Anp Agencia Nacional Do Petroleo Gas BR Consolidada (07!12!2021)Document33 paginiResolucao 828 2020 Anp Agencia Nacional Do Petroleo Gas BR Consolidada (07!12!2021)José EstevesÎncă nu există evaluări
- Pasta QuímicaDocument12 paginiPasta QuímicaFrancisco Nunes NegreirosÎncă nu există evaluări
- Tendências e Desafios de Espumas À Base de Amido para Uso Como Embalagem de Alimentos e Recipiente para AlimentosDocument51 paginiTendências e Desafios de Espumas À Base de Amido para Uso Como Embalagem de Alimentos e Recipiente para AlimentosGabriela PozzebonÎncă nu există evaluări
- GPT 2020 It Digeob 06 Simbolos e Abreviaturas El Quimicos Minerais e Rochas 01 07 2020Document13 paginiGPT 2020 It Digeob 06 Simbolos e Abreviaturas El Quimicos Minerais e Rochas 01 07 2020Ronaldo PereiraÎncă nu există evaluări
- Defesas ManíacasDocument6 paginiDefesas Maníacasmariana luz tavaresÎncă nu există evaluări
- Bula MinilaxDocument6 paginiBula MinilaxEllen AraujoÎncă nu există evaluări
- Avaliação Do Risco de SuicídioDocument52 paginiAvaliação Do Risco de SuicídioDaniela Paes Landim BorgesÎncă nu există evaluări
- Certificados de Calidad MaterialesDocument12 paginiCertificados de Calidad MaterialesLaura CondizaÎncă nu există evaluări
- Aula 03 Semana Da BioquimicaDocument14 paginiAula 03 Semana Da Bioquimicaandre almeidaÎncă nu există evaluări
- DownloadDocument8 paginiDownloadIsabella MariaÎncă nu există evaluări