S-ar putea să vă placă și
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Origami Car (VW Bug)Document5 paginiOrigami Car (VW Bug)Jesse GreerÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Science Volume 324 Issue 5923 2009 (Doi 10.1126/science.324.5923.28) Pennisi, E. - OrIGINS - On The Origin of Flowering PlantsDocument4 paginiScience Volume 324 Issue 5923 2009 (Doi 10.1126/science.324.5923.28) Pennisi, E. - OrIGINS - On The Origin of Flowering PlantsDiana MendoncaÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5795)
- Praying Mantis Robert LangDocument7 paginiPraying Mantis Robert LangJae ElkindÎncă nu există evaluări
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Yoda JediMaster PDFDocument7 paginiYoda JediMaster PDFDiana MendoncaÎncă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Grana: Striate Pollen: New Early Eudicots FromDocument22 paginiGrana: Striate Pollen: New Early Eudicots FromDiana MendoncaÎncă nu există evaluări
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- Peter K Endress: Evolution of Floral SymmetryDocument6 paginiPeter K Endress: Evolution of Floral SymmetryDiana MendoncaÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Zoological Journal of The Linnean Society Volume 45 Issue 304 1964 (Doi 10.1111/j.1096-3642.1964.Tb00488.x) Robert L. Carroll - The Earliest ReptilesDocument24 paginiZoological Journal of The Linnean Society Volume 45 Issue 304 1964 (Doi 10.1111/j.1096-3642.1964.Tb00488.x) Robert L. Carroll - The Earliest ReptilesDiana MendoncaÎncă nu există evaluări
- The Origin and Evolution of The Angiosperm Carpel: David Winship Taylor and Gretchen KirchnerDocument25 paginiThe Origin and Evolution of The Angiosperm Carpel: David Winship Taylor and Gretchen KirchnerDiana MendoncaÎncă nu există evaluări
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Geobios Volume 28 Issue 5 1995 (Doi 10.1016/s0016-6995 (95) 80211-8) Blanka Pacltova - The Enigma of Angiosperm Origins PDFDocument1 paginăGeobios Volume 28 Issue 5 1995 (Doi 10.1016/s0016-6995 (95) 80211-8) Blanka Pacltova - The Enigma of Angiosperm Origins PDFDiana MendoncaÎncă nu există evaluări
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Beaded GardenDocument122 paginiThe Beaded GardenDiana MendoncaÎncă nu există evaluări
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Companion Animals: 2. Background To Stray Animal IssuesDocument16 paginiCompanion Animals: 2. Background To Stray Animal IssuesDiana MendoncaÎncă nu există evaluări
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Kingdom Animalia Deep FlowchartDocument2 paginiKingdom Animalia Deep Flowchartmeerab uroojÎncă nu există evaluări
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Soal Pat Genap Kelas Viii Bahasa Inggris Tapel 2020-2021 Foskam - 2Document4 paginiSoal Pat Genap Kelas Viii Bahasa Inggris Tapel 2020-2021 Foskam - 2nazar azkhafÎncă nu există evaluări
- Denver Zoo Masterplan SummaryDocument22 paginiDenver Zoo Masterplan SummaryThe Denver postÎncă nu există evaluări
- ZP MPDocument2 paginiZP MPNagbhushan_cmÎncă nu există evaluări
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Wildlife Fact File - Birds - Pgs. 231-240Document20 paginiWildlife Fact File - Birds - Pgs. 231-240ClearMind84100% (1)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Avise, 2004. Molecular Markers, Natural History, and EvolutionDocument667 paginiAvise, 2004. Molecular Markers, Natural History, and EvolutionKaio BarrosÎncă nu există evaluări
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Chapter 3 Biology Form 4Document37 paginiChapter 3 Biology Form 4CuteOnion Furmon100% (1)
- Dissecting A Whole ChickenDocument5 paginiDissecting A Whole Chickenbrianmore10Încă nu există evaluări
- Darwins Ikthiologi PDFDocument367 paginiDarwins Ikthiologi PDFcaoryjunÎncă nu există evaluări
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- BIOLAB FROG Organ HistologyDocument7 paginiBIOLAB FROG Organ HistologyCloie Anne Rabinetas100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1091)
- Introduction To Cladistics.: Character A-Willow B-Parrot C-CowDocument4 paginiIntroduction To Cladistics.: Character A-Willow B-Parrot C-Cowskline3Încă nu există evaluări
- Reading and WritingDocument1 paginăReading and Writingkreigblitz9801Încă nu există evaluări
- 1970 - Lewontin, R. - The Units of Selection PDFDocument19 pagini1970 - Lewontin, R. - The Units of Selection PDFvltg_35643Încă nu există evaluări
- ICMR JRF 2015 Previous - Model Question Paper - EasybiologyclassDocument10 paginiICMR JRF 2015 Previous - Model Question Paper - EasybiologyclassSharad GajuryalÎncă nu există evaluări
- Pro To Zoology 1954 KudoDocument988 paginiPro To Zoology 1954 KudoClaau GomezÎncă nu există evaluări
- Human EvolutionDocument23 paginiHuman EvolutionArpit Shukla100% (2)
- The Fox Plays The Bear A Trick: Ion CreangaDocument20 paginiThe Fox Plays The Bear A Trick: Ion CreangaChaitanya K SÎncă nu există evaluări
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Anatomical Landmarks - MaxillaDocument5 paginiAnatomical Landmarks - MaxillaAdarsh A SÎncă nu există evaluări
- Posterior Palatal SealDocument61 paginiPosterior Palatal SealsmritinarayanÎncă nu există evaluări
- PilandokDocument4 paginiPilandokpatricksvdÎncă nu există evaluări
- Dichotomous Key ExerciseDocument64 paginiDichotomous Key ExerciseAnne Sharmaine OchoaÎncă nu există evaluări
- External Anatomy of The FrogDocument19 paginiExternal Anatomy of The Frogabigail_estrecho83% (6)
- RBSC Art-Falconry Casey-Wood SK321F87Document767 paginiRBSC Art-Falconry Casey-Wood SK321F87Branko NikolicÎncă nu există evaluări
- Butterflies W JavaDocument26 paginiButterflies W JavaMauLidia Achmad Rukun Marmy100% (4)
- 102PDFDocument41 pagini102PDFmayorca2310Încă nu există evaluări
- Florence: Series - 1BDocument20 paginiFlorence: Series - 1BKannan GamingÎncă nu există evaluări
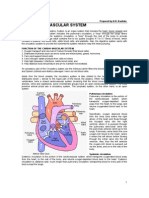
- Cardiovascular SystemDocument5 paginiCardiovascular SystemAnonymous HXLczq3100% (1)
- Courtship BehaviourDocument36 paginiCourtship BehaviourMunzareen RiazÎncă nu există evaluări
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Operator Position, Visibility, Illumination, and Finger RestDocument14 paginiOperator Position, Visibility, Illumination, and Finger RestjihanÎncă nu există evaluări
- Lec No 8Document5 paginiLec No 8Pratibha ChÎncă nu există evaluări