S-ar putea să vă placă și
- Fruit Salad 115 RecipeDocument1 paginăFruit Salad 115 RecipeEricka GalangÎncă nu există evaluări
- Research of The WeekDocument4 paginiResearch of The WeekEricka GalangÎncă nu există evaluări
- DNA Extraction and CharacterizationDocument4 paginiDNA Extraction and CharacterizationEricka Galang100% (1)
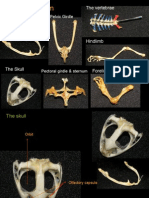
- Cat MusclesDocument15 paginiCat MusclesEricka GalangÎncă nu există evaluări
- Banmal ChordsDocument2 paginiBanmal ChordsMaria ConnerÎncă nu există evaluări
- Reaction PaperDocument10 paginiReaction PaperEricka Galang100% (1)
- Enzyme KineticsDocument3 paginiEnzyme KineticsEricka GalangÎncă nu există evaluări
- On The Job Reaction PaperDocument2 paginiOn The Job Reaction PaperEricka Galang88% (8)
- Characterization of Proteins Using Ion Exchange Chromatography and Gel Filtration ChromatographyDocument4 paginiCharacterization of Proteins Using Ion Exchange Chromatography and Gel Filtration ChromatographyEricka GalangÎncă nu există evaluări
- Frog Germ LayerDocument4 paginiFrog Germ LayerEricka GalangÎncă nu există evaluări
- Nucleophilic SubstitutionDocument8 paginiNucleophilic SubstitutionEricka GalangÎncă nu există evaluări
- ProtozoaDocument1 paginăProtozoaEricka GalangÎncă nu există evaluări
- Synthesis of An Alkyl HalidesDocument3 paginiSynthesis of An Alkyl HalidesEricka GalangÎncă nu există evaluări
- 6.5a Strong and Weak Acids and BasesDocument14 pagini6.5a Strong and Weak Acids and BasesEricka GalangÎncă nu există evaluări
- LE 3 NotesDocument13 paginiLE 3 NotesEricka GalangÎncă nu există evaluări
- ALGAEDocument1 paginăALGAEEricka GalangÎncă nu există evaluări
- Synthesis of An Alkyl HalideDocument1 paginăSynthesis of An Alkyl HalideEricka GalangÎncă nu există evaluări
- MSDS For Experiment 6 of Chem 31.1 (Synthesis of An Alkyl Halide)Document3 paginiMSDS For Experiment 6 of Chem 31.1 (Synthesis of An Alkyl Halide)Ericka GalangÎncă nu există evaluări
- From Musculature and Skeletal SystemDocument23 paginiFrom Musculature and Skeletal SystemEricka GalangÎncă nu există evaluări
- Leadership Report About South East Asia For SEA30Document4 paginiLeadership Report About South East Asia For SEA30Ericka GalangÎncă nu există evaluări
- Chapter 15Document40 paginiChapter 15Ericka GalangÎncă nu există evaluări
- Chem 26.1 Experiment 6 Answers To QuestionsDocument2 paginiChem 26.1 Experiment 6 Answers To QuestionsEricka GalangÎncă nu există evaluări
- Chem 26.1 Experiment 3 The Iodine Clock Reactions Formal ReportDocument5 paginiChem 26.1 Experiment 3 The Iodine Clock Reactions Formal ReportEricka Galang100% (1)
- IncentiveDocument1 paginăIncentiveEricka GalangÎncă nu există evaluări
- RDR 1Document2 paginiRDR 1Ericka GalangÎncă nu există evaluări
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Tau-Targeting Antisense Oligonucleotide Mapt in Mild Alzheimer's Disease: A Phase 1b, Randomized, Placebo-Controlled TrialDocument20 paginiTau-Targeting Antisense Oligonucleotide Mapt in Mild Alzheimer's Disease: A Phase 1b, Randomized, Placebo-Controlled TrialSiddharth NarsipurÎncă nu există evaluări
- The RH Negative Factor BookDocument117 paginiThe RH Negative Factor BookRoberta Ruth Hill67% (3)
- 269 - Embryology Physiology) Development of The Heart  - ÏDocument11 pagini269 - Embryology Physiology) Development of The Heart  - ÏFood Safety CommunityÎncă nu există evaluări
- Hyperparathyrodism Hypoparathyrodism: Dr. A. Wahyudi Pababbari, SPPD, FinasimDocument48 paginiHyperparathyrodism Hypoparathyrodism: Dr. A. Wahyudi Pababbari, SPPD, FinasimKahfiyahÎncă nu există evaluări
- AutoimunDocument23 paginiAutoimunEllya AfianiÎncă nu există evaluări
- ABMLI Sample Questions 000Document7 paginiABMLI Sample Questions 000samy100% (1)
- Popova 2016Document78 paginiPopova 2016nenymurniatiÎncă nu există evaluări
- Anaphylactic ShockDocument29 paginiAnaphylactic ShockHIND100% (7)
- Paper Pet ProjectDocument27 paginiPaper Pet Projectapi-406104878Încă nu există evaluări
- Lymphomas: Varun. M M-Pharm (Pharmacology) IyriisemDocument32 paginiLymphomas: Varun. M M-Pharm (Pharmacology) IyriisemPintoo Pintu100% (2)
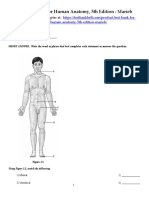
- Test Bank For Human Anatomy 5th Edition MariebDocument18 paginiTest Bank For Human Anatomy 5th Edition MariebSandra Brown100% (5)
- Biology SL P2Document8 paginiBiology SL P2KenanÎncă nu există evaluări
- Referensi CK2Document1 paginăReferensi CK2Rahma UlfaÎncă nu există evaluări
- Swyer SyndromeDocument1 paginăSwyer SyndromeHaÎncă nu există evaluări
- 9q Deletions Including 9q33 FTNPDocument10 pagini9q Deletions Including 9q33 FTNPMartin GirardÎncă nu există evaluări
- Addiction AND Upregulation BUISSON 2002Document7 paginiAddiction AND Upregulation BUISSON 2002Andra SilvaÎncă nu există evaluări
- Genetic Influences in Human BehaviorDocument24 paginiGenetic Influences in Human BehaviorGuillermo ArriagaÎncă nu există evaluări
- Icsa Usmle Courses ContentDocument18 paginiIcsa Usmle Courses ContentdostoviskyÎncă nu există evaluări
- Chemical Carcinogenesis I and Ii: Michael Lea 2016Document31 paginiChemical Carcinogenesis I and Ii: Michael Lea 2016aasiyaÎncă nu există evaluări
- Full Paper AIC 2018Document28 paginiFull Paper AIC 2018yantiÎncă nu există evaluări
- Medical Genetics SummariesDocument462 paginiMedical Genetics SummariesMohd RahimiÎncă nu există evaluări
- Exchange Blood Transfusion 2Document15 paginiExchange Blood Transfusion 2Sarah100% (1)
- Tife Chapter Two-2Document37 paginiTife Chapter Two-2Ajegbomogun SundayÎncă nu există evaluări
- CauliflowerDocument4 paginiCauliflowerIzhar Ahmad DanishÎncă nu există evaluări
- Diagnosis Power PointDocument26 paginiDiagnosis Power PointGlenn ChavezÎncă nu există evaluări
- Purupura Henoch Schonlein Tipe Bula HemoragikDocument51 paginiPurupura Henoch Schonlein Tipe Bula HemoragikMedina MarwanÎncă nu există evaluări
- Advanced Prostate Cancer: Treatment Advances and Future DirectionsDocument14 paginiAdvanced Prostate Cancer: Treatment Advances and Future DirectionsMishel Rodriguez GuzmanÎncă nu există evaluări
- B5 Lecture Slides (CMF) Folate AntagonistsDocument15 paginiB5 Lecture Slides (CMF) Folate AntagonistsZhen Yuan GaryÎncă nu există evaluări
- Tura KuDocument104 paginiTura KuFitzgerald BillyÎncă nu există evaluări
- Mechanism of HypertensionDocument4 paginiMechanism of HypertensionAlya Putri KhairaniÎncă nu există evaluări