S-ar putea să vă placă și
- Krebs CycleDocument2 paginiKrebs CycleSmalldaniel0% (3)
- North Carolina Biology EOC Study GuideDocument38 paginiNorth Carolina Biology EOC Study GuideRaquel Galdamez-GaldamezÎncă nu există evaluări
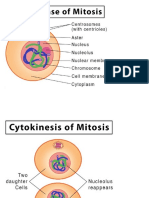
- Mitosis Cell Division PDFDocument8 paginiMitosis Cell Division PDFPerry SinÎncă nu există evaluări
- Cell Cycle N Cell DivisionDocument3 paginiCell Cycle N Cell DivisionDanish UllahÎncă nu există evaluări
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceDe la EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceÎncă nu există evaluări
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsDe la EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsEvaluare: 5 din 5 stele5/5 (1)
- Structure of DNA: Teacher Notes and AnswersDocument4 paginiStructure of DNA: Teacher Notes and Answersyawahab100% (1)
- The Cell Cycle-TRFDocument8 paginiThe Cell Cycle-TRFmaniiiiiiiiÎncă nu există evaluări
- Referat asma-PARUDocument26 paginiReferat asma-PARUPutri Andansari100% (1)
- Cell Cycle Stages & PhasesDocument11 paginiCell Cycle Stages & PhasesJessie Katrisha TayagÎncă nu există evaluări
- Cell Respiration: ATP from GlucoseDocument11 paginiCell Respiration: ATP from GlucoseNiña Jennifer Potter Vergara100% (2)
- Chapter 9 Cell Cycle Chromosome and Mitosis 2018Document42 paginiChapter 9 Cell Cycle Chromosome and Mitosis 2018Nehjoy GerangueÎncă nu există evaluări
- TEST BANK - Chapter 3 - Amino Acids, Peptides, and ProteinsDocument31 paginiTEST BANK - Chapter 3 - Amino Acids, Peptides, and ProteinsAndreaÎncă nu există evaluări
- r7412311 Metabolic EngineeringDocument4 paginir7412311 Metabolic EngineeringsivabharathamurthyÎncă nu există evaluări
- Cell Division ProcessDocument30 paginiCell Division ProcessM Noaman AkbarÎncă nu există evaluări
- Interphase Prophase: ChromoDocument12 paginiInterphase Prophase: ChromoNkosi JupiterÎncă nu există evaluări
- MITOSIS and MEIOSISDocument26 paginiMITOSIS and MEIOSISMuskan ShreeÎncă nu există evaluări
- Cell DivisionDocument11 paginiCell DivisionDr Nicholas AdeÎncă nu există evaluări
- Biology AssgDocument19 paginiBiology AssgHABTEMARIAM ERTBANÎncă nu există evaluări
- MITOSISDocument5 paginiMITOSISdeepaÎncă nu există evaluări
- Cell Cycle: Mitosis and MeiosisDocument10 paginiCell Cycle: Mitosis and MeiosisdeepaÎncă nu există evaluări
- Chromosomes and Cell ReproductionDocument29 paginiChromosomes and Cell ReproductionTahreem BhattiÎncă nu există evaluări
- Cell DivisionDocument32 paginiCell DivisionRhea GulayÎncă nu există evaluări
- Zoology Lecture 5Document7 paginiZoology Lecture 5Stephen OcheaÎncă nu există evaluări
- MitosisDocument7 paginiMitosisRishikesh BhintadeÎncă nu există evaluări
- Unit 3 NotesDocument6 paginiUnit 3 NotesibtiÎncă nu există evaluări
- Chapter 9 Cell DivisionDocument4 paginiChapter 9 Cell DivisionMaria Jose UribeÎncă nu există evaluări
- Biotechnology NotesDocument21 paginiBiotechnology NotesRommel BauzaÎncă nu există evaluări
- The Stages and Purpose of MitosisDocument9 paginiThe Stages and Purpose of MitosisJeazel MosendoÎncă nu există evaluări
- 2.5 - Cell DivisionDocument5 pagini2.5 - Cell DivisionAlvin JovanÎncă nu există evaluări
- Mitosis: Aarifa Hoosein-RiyasatDocument28 paginiMitosis: Aarifa Hoosein-RiyasatAkiela JohnsonÎncă nu există evaluări
- Genetics: Cell Cycle Mitosis MeiosisDocument60 paginiGenetics: Cell Cycle Mitosis MeiosisPatches24 Patches24Încă nu există evaluări
- 4.2 Cell As A Unit of LifeDocument7 pagini4.2 Cell As A Unit of Lifehola adiosÎncă nu există evaluări
- 5.cell DivisionDocument54 pagini5.cell DivisionRaymond LundaÎncă nu există evaluări
- Meiosis and Mitosis: The Cell Division ProcessesDocument17 paginiMeiosis and Mitosis: The Cell Division ProcessesMuskan ShreeÎncă nu există evaluări
- Cell CycleDocument111 paginiCell Cyclepreyaaa pascuaÎncă nu există evaluări
- CITOSISDocument3 paginiCITOSISKylliíanÎncă nu există evaluări
- MitosisDocument3 paginiMitosisLea Mari JavierÎncă nu există evaluări
- Degree Assignment SolutionDocument4 paginiDegree Assignment SolutionESTHER MWEEMBAÎncă nu există evaluări
- 1.3 - Mitosis-1Document27 pagini1.3 - Mitosis-1sophiewayman07Încă nu există evaluări
- Cell Division MitosisDocument6 paginiCell Division MitosisIroldÎncă nu există evaluări
- Cell Division MitosisDocument6 paginiCell Division MitosisIroldÎncă nu există evaluări
- Copia de MITOSIS Texts FotoDocument2 paginiCopia de MITOSIS Texts FotoMarcos Nuñez CarreteroÎncă nu există evaluări
- Cell Cycle and MitosisDocument49 paginiCell Cycle and Mitosishazharomar958Încă nu există evaluări
- Cell CycleDocument19 paginiCell CycleMoffat KaprezzoÎncă nu există evaluări
- Cell Division StagesDocument118 paginiCell Division StagesNora AfidaÎncă nu există evaluări
- Mitosis and MeiosisDocument13 paginiMitosis and MeiosisS3 Oxibillo, Gabriel Dominic C.Încă nu există evaluări
- MODULE 3 - MitosisDocument3 paginiMODULE 3 - MitosisNicoleBenaguaÎncă nu există evaluări
- Mitosis Divides Cell Nucleus ChromosomesDocument3 paginiMitosis Divides Cell Nucleus ChromosomesVinodh RavindaranÎncă nu există evaluări
- Cell DivisionDocument28 paginiCell DivisionPalagiri MadhuÎncă nu există evaluări
- CH 15 NotesDocument5 paginiCH 15 NotesCJÎncă nu există evaluări
- Cell Division and Meiosis ExplainedDocument92 paginiCell Division and Meiosis ExplainedHarpreet KaurÎncă nu există evaluări
- MitosisDocument3 paginiMitosisSabina MaharjanÎncă nu există evaluări
- The Cell CycleDocument9 paginiThe Cell Cyclesalahuddin_md5935Încă nu există evaluări
- Lesson 7 - MitosisDocument5 paginiLesson 7 - MitosisReuben Al'jaldiÎncă nu există evaluări
- Agric QuizDocument3 paginiAgric QuizKelsy SalazarÎncă nu există evaluări
- Mitosis Meiosis NotesDocument5 paginiMitosis Meiosis NotesnicoleÎncă nu există evaluări
- Cell Division: Growth, Replication and DistributionDocument3 paginiCell Division: Growth, Replication and DistributionMohnish MÎncă nu există evaluări
- Lecture 7 - 23.12.2022Document30 paginiLecture 7 - 23.12.2022Adnan Mohammad Adnan HailatÎncă nu există evaluări
- Cell CycleDocument6 paginiCell Cyclewebpixel servicesÎncă nu există evaluări
- Mitosis: The Process by Which Cells DivideDocument7 paginiMitosis: The Process by Which Cells DivideLeo Paolo LuceroÎncă nu există evaluări
- CytokinesisDocument15 paginiCytokinesisMar Delos ReyesÎncă nu există evaluări
- Biosciweek6 11Document43 paginiBiosciweek6 11Jesalyn AbrinicaÎncă nu există evaluări
- Folio Kerja Kursus Biology SPMDocument29 paginiFolio Kerja Kursus Biology SPMzam_ir100% (2)
- The Cell Cycle (Mitosis)Document2 paginiThe Cell Cycle (Mitosis)magdalenasilviÎncă nu există evaluări
- ApbiopopulationgeneticsDocument17 paginiApbiopopulationgeneticsюрий локтионовÎncă nu există evaluări
- ApbiopopulationgeneticsDocument17 paginiApbiopopulationgeneticsюрий локтионовÎncă nu există evaluări
- Receitas 01Document1 paginăReceitas 01Adriana de CarvalhoÎncă nu există evaluări
- Ya-DongDocument1 paginăYa-Dongюрий локтионовÎncă nu există evaluări
- Receitas 01Document1 paginăReceitas 01Adriana de CarvalhoÎncă nu există evaluări
- Volkswagen S ComebackDocument1 paginăVolkswagen S Comebackюрий локтионовÎncă nu există evaluări
- Smartshare PC SW (Dlna)Document1 paginăSmartshare PC SW (Dlna)jr100100Încă nu există evaluări
- EmailDocument1 paginăEmailюрий локтионовÎncă nu există evaluări
- Receitas 01Document1 paginăReceitas 01Adriana de CarvalhoÎncă nu există evaluări
- Smartshare PC SW (Dlna)Document1 paginăSmartshare PC SW (Dlna)jr100100Încă nu există evaluări
- Receitas 01Document1 paginăReceitas 01Adriana de CarvalhoÎncă nu există evaluări
- Active Work Snapshot Wasabi Ventures 3 4 2015 PDFDocument2 paginiActive Work Snapshot Wasabi Ventures 3 4 2015 PDFюрий локтионовÎncă nu există evaluări
- Russell-AthleticDocument28 paginiRussell-Athleticюрий локтионовÎncă nu există evaluări
- Walmart S Global StrategiesDocument2 paginiWalmart S Global Strategiesюрий локтионовÎncă nu există evaluări
- Motivation Across CulturesDocument2 paginiMotivation Across Culturesюрий локтионовÎncă nu există evaluări
- RocheDocument1 paginăRocheюрий локтионовÎncă nu există evaluări
- Interconnected-WorldDocument1 paginăInterconnected-Worldюрий локтионовÎncă nu există evaluări
- RussiaDocument1 paginăRussiaюрий локтионовÎncă nu există evaluări
- India Vs America CommunicationDocument1 paginăIndia Vs America Communicationюрий локтионовÎncă nu există evaluări
- Importance of Goal SettingDocument1 paginăImportance of Goal Settingюрий локтионовÎncă nu există evaluări
- Chapter-14Document1 paginăChapter-14юрий локтионовÎncă nu există evaluări
- HSBC in ChinaDocument3 paginiHSBC in Chinaюрий локтионов100% (1)
- Femicide-MemoDocument2 paginiFemicide-Memoюрий локтионовÎncă nu există evaluări
- China S MillenialsDocument1 paginăChina S Millenialsюрий локтионовÎncă nu există evaluări
- Jelin-MemoDocument1 paginăJelin-Memoюрий локтионовÎncă nu există evaluări
- Afghanistan Info SheetDocument4 paginiAfghanistan Info Sheetюрий локтионовÎncă nu există evaluări
- Big Pharma Goes GlobalDocument1 paginăBig Pharma Goes Globalюрий локтионовÎncă nu există evaluări
- Extra-CreditDocument1 paginăExtra-Creditюрий локтионовÎncă nu există evaluări
- Bolender-MemoDocument2 paginiBolender-Memoюрий локтионовÎncă nu există evaluări
- Bandits Article NotesDocument2 paginiBandits Article Notesюрий локтионовÎncă nu există evaluări
- Chapter 5 QuestionDocument26 paginiChapter 5 QuestionNurul AzuwinÎncă nu există evaluări
- Meiosis Notes MsDocument28 paginiMeiosis Notes Msapi-44119921Încă nu există evaluări
- PER. 5 Enzymes in Genetics Engineering - 2Document38 paginiPER. 5 Enzymes in Genetics Engineering - 2sri wahyuniÎncă nu există evaluări
- Genomics and Bioinformatics: Peter Gregory and Senthil NatesanDocument22 paginiGenomics and Bioinformatics: Peter Gregory and Senthil NatesanajaybioinfoÎncă nu există evaluări
- UBC Genetics 234 Fundamentals CourseDocument2 paginiUBC Genetics 234 Fundamentals CourseJustin ChauÎncă nu există evaluări
- J. Biol. Chem.-1988-Meister-17205-8Document4 paginiJ. Biol. Chem.-1988-Meister-17205-8Pepé DubÎncă nu există evaluări
- Microbiology Jeopardy ReviewDocument31 paginiMicrobiology Jeopardy Reviewjmiller623Încă nu există evaluări
- Serological cross-reactivity-HLA-A Locus: 1 CregDocument5 paginiSerological cross-reactivity-HLA-A Locus: 1 CregGeorgia MarcussoÎncă nu există evaluări
- Chapter 14 Lecture Notes: Nucleic Acids: Educational GoalsDocument28 paginiChapter 14 Lecture Notes: Nucleic Acids: Educational GoalsMehran AhmadÎncă nu există evaluări
- PCR DNA Fingerprinting Forensic AnalysisDocument9 paginiPCR DNA Fingerprinting Forensic Analysiskhanyiso kalyataÎncă nu există evaluări
- Ratibrom 2Document2 paginiRatibrom 2Frontline0712Încă nu există evaluări
- CSIR NET Life Sciences Previous Year Solution June 2022 S1 Part BDocument56 paginiCSIR NET Life Sciences Previous Year Solution June 2022 S1 Part Bpiyush raghavÎncă nu există evaluări
- History of DNA and the Central DogmaDocument1 paginăHistory of DNA and the Central Dogmaemarie8panerioÎncă nu există evaluări
- Cloning Blunt-End DNA Fragments Into The Pgem - T Vector SystemsDocument5 paginiCloning Blunt-End DNA Fragments Into The Pgem - T Vector SystemsKhanh NhuÎncă nu există evaluări
- Mid Question Bank (Proposed)Document3 paginiMid Question Bank (Proposed)Zahidul HassanÎncă nu există evaluări
- M.Sc. - Molecular Human GeneticsDocument32 paginiM.Sc. - Molecular Human GeneticsSiman SimanÎncă nu există evaluări
- Gateway Cloning ManualDocument63 paginiGateway Cloning ManualdavidjkimÎncă nu există evaluări
- Chapter 1 Cell StructureDocument37 paginiChapter 1 Cell StructureNawafÎncă nu există evaluări
- Question Report 45 PDFDocument10 paginiQuestion Report 45 PDFHorrid HenryÎncă nu există evaluări
- Cell Structure and Funtions 2 PDFDocument49 paginiCell Structure and Funtions 2 PDFEllie MarrisÎncă nu există evaluări
- MeiosisDocument10 paginiMeiosisNoor Hafezah Mohd MokhtiarÎncă nu există evaluări
- Blue WhiteDocument2 paginiBlue WhiteShyamala RamalingamÎncă nu există evaluări
- Assessment of DNA Yield and Purity An Overlooked DDocument6 paginiAssessment of DNA Yield and Purity An Overlooked DKatrinaÎncă nu există evaluări