S-ar putea să vă placă și
- Embryogenesis: Dicots, Monocots, and GymnospermsDocument61 paginiEmbryogenesis: Dicots, Monocots, and GymnospermsMae ChaÎncă nu există evaluări
- 10 EmbryogenesisDocument14 pagini10 EmbryogenesisPhenyo MmerekiÎncă nu există evaluări
- Membrane Models: Gorter and Grendel's Membrane Theory (1920)Document5 paginiMembrane Models: Gorter and Grendel's Membrane Theory (1920)Hafiz AhmadÎncă nu există evaluări
- PhotoperiodismDocument14 paginiPhotoperiodismArun Sreenivasan IyengarÎncă nu există evaluări
- Plant Embryo DevelopmentDocument29 paginiPlant Embryo DevelopmentMa-anJaneDiamos100% (1)
- Plant KingdomDocument16 paginiPlant Kingdomshriyansh singhaniaÎncă nu există evaluări
- History of ZoologyDocument4 paginiHistory of ZoologyBen Gabriel MaghuyopÎncă nu există evaluări
- 36 - ch2 - Reproduction in PlantsDocument21 pagini36 - ch2 - Reproduction in PlantsAnonymous Azxx3Kp9100% (1)
- Eukaryotic Gene Regulation NotesDocument16 paginiEukaryotic Gene Regulation Notesapi-277471896Încă nu există evaluări
- Biology PDFDocument344 paginiBiology PDFUrmilaAnantÎncă nu există evaluări
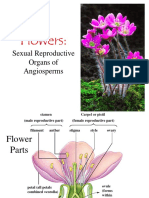
- Flowers:: Sexual Reproductive Organs of AngiospermsDocument106 paginiFlowers:: Sexual Reproductive Organs of AngiospermsAlfonso PlantillaÎncă nu există evaluări
- Cell Cycle Cell Division PDFDocument13 paginiCell Cycle Cell Division PDFmuzamil shabir100% (1)
- Inhibitory Effects of Ribbon Sea Lettuce (Ulva Reticulata) Extracts Against Klebsiella PneumoniaeDocument45 paginiInhibitory Effects of Ribbon Sea Lettuce (Ulva Reticulata) Extracts Against Klebsiella PneumoniaeVanessa Marie MatutesÎncă nu există evaluări
- Laboratory Exercise 1A - Yarn ModelsDocument2 paginiLaboratory Exercise 1A - Yarn ModelsGianjim HubillaÎncă nu există evaluări
- Plants Reproduction and DevelopmentDocument6 paginiPlants Reproduction and DevelopmentAlwìn GarciaÎncă nu există evaluări
- Network Structure in Root, Stem, and LeafDocument5 paginiNetwork Structure in Root, Stem, and LeafMarwana SuaibÎncă nu există evaluări
- Plant Taxonomy NotesDocument38 paginiPlant Taxonomy Notesopolla nianorÎncă nu există evaluări
- 5 Male Sterility 29-3-17Document64 pagini5 Male Sterility 29-3-17Aizaz AliÎncă nu există evaluări
- Mitosis and Meiosis: Cell DivisionDocument36 paginiMitosis and Meiosis: Cell DivisionHeart VidalÎncă nu există evaluări
- 4 Animal KingdomDocument33 pagini4 Animal KingdomS SÎncă nu există evaluări
- Respiratory System: StructureDocument29 paginiRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Exercise 1 The Cell 1Document6 paginiExercise 1 The Cell 1MJMadlangbayan100% (1)
- MeiosisDocument8 paginiMeiosisJohn Chiyu Garde-Labite Azuki100% (1)
- M.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarDocument39 paginiM.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarM.PRASAD NAIDUÎncă nu există evaluări
- Class: 8 Science Chapter - 8 Cell - Structure and FunctionsDocument10 paginiClass: 8 Science Chapter - 8 Cell - Structure and FunctionsAmiteshwar SinghÎncă nu există evaluări
- Unit 13 Cleavage and GastrulationDocument26 paginiUnit 13 Cleavage and GastrulationFERHAEEZA KALAYAKANÎncă nu există evaluări
- Sex Determination in DrosophilaDocument19 paginiSex Determination in DrosophilaSheerin Sulthana100% (2)
- MarchantiophytaDocument9 paginiMarchantiophytax456456456xÎncă nu există evaluări
- Approval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Document12 paginiApproval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Lhely Andi NurlaeliÎncă nu există evaluări
- Lab Report 1Document6 paginiLab Report 1Margarita Kotsev100% (1)
- Unit-4 4. Chemistry CarbohydratesDocument45 paginiUnit-4 4. Chemistry CarbohydratesKiya Alemu100% (1)
- PHCG 201 CytologyDocument113 paginiPHCG 201 CytologyIbn HaarūnÎncă nu există evaluări
- Plant CellDocument112 paginiPlant CellAlfonso PlantillaÎncă nu există evaluări
- Cell DifferentiationDocument16 paginiCell DifferentiationHarika KeshavÎncă nu există evaluări
- Cell+ +Cell+MembraneDocument26 paginiCell+ +Cell+Membraneemancipation1506Încă nu există evaluări
- Dihybrid ProblemDocument5 paginiDihybrid ProblemJohn Keir Pedregosa PasquinÎncă nu există evaluări
- 5.3 Understanding Genes and ChromosomesDocument11 pagini5.3 Understanding Genes and ChromosomesNoor Hidayah SambliÎncă nu există evaluări
- Lab 1 Plant MorphologyDocument5 paginiLab 1 Plant MorphologyNabilah SyahirahÎncă nu există evaluări
- Bryophytes: Dr. Rukhshana ParveenDocument7 paginiBryophytes: Dr. Rukhshana ParveenSakshi SharmaÎncă nu există evaluări
- Biol-111 - Mendelian Genetics WorksheetDocument10 paginiBiol-111 - Mendelian Genetics WorksheetKing JoshÎncă nu există evaluări
- Theories of Organic EvolutionDocument15 paginiTheories of Organic EvolutionAdityaÎncă nu există evaluări
- Evolution Review Packet Key EditableDocument4 paginiEvolution Review Packet Key EditableMiljoy Delegado100% (1)
- BiotechnologyDocument18 paginiBiotechnologyZanib SarfrazÎncă nu există evaluări
- Syllabus in Chemical Biology III (Biomolecules) (TCB3)Document12 paginiSyllabus in Chemical Biology III (Biomolecules) (TCB3)Yuri PaderesÎncă nu există evaluări
- Chloroplast GenomeDocument13 paginiChloroplast Genomelordniklaus100% (2)
- Basic Concept of Alage - Bryophyte - Pteridophyte & GymnospermDocument19 paginiBasic Concept of Alage - Bryophyte - Pteridophyte & GymnospermDivyansha Sharma100% (1)
- Osmosis, Diffusion, Active TransportDocument21 paginiOsmosis, Diffusion, Active TransportPrince SanjiÎncă nu există evaluări
- 11.2 Domain Bacteria and ArchaeaDocument39 pagini11.2 Domain Bacteria and ArchaeaZkdlin SpaceÎncă nu există evaluări
- SUMMURIZEDocument55 paginiSUMMURIZEWaren LlorenÎncă nu există evaluări
- Transposons in HumansDocument22 paginiTransposons in HumansRishiÎncă nu există evaluări
- Cell Bio Assignment - Plasma Membrane and Basement MembraneDocument2 paginiCell Bio Assignment - Plasma Membrane and Basement Membranerahul mauryaÎncă nu există evaluări
- Cell Biology Practical 3 PDFDocument11 paginiCell Biology Practical 3 PDFEswaran Singam100% (1)
- 1 Pteridophyta PsilotopsidaDocument25 pagini1 Pteridophyta PsilotopsidaCDB 1st Semester 2077100% (1)
- BIO101 Lec 04-DNA Denaturation and RenaturationDocument29 paginiBIO101 Lec 04-DNA Denaturation and RenaturationMika MaravillaÎncă nu există evaluări
- Handouts-Lect. 29 - Introduction To Kingdom Plantae PDFDocument6 paginiHandouts-Lect. 29 - Introduction To Kingdom Plantae PDFRizwan BalochÎncă nu există evaluări
- Gnetales PG-I SushilkrsinghDocument22 paginiGnetales PG-I SushilkrsinghN Ganapathi Kumar100% (1)
- Eukaryotic Cell OrganellesDocument7 paginiEukaryotic Cell OrganellesMary100% (2)
- BryophytesDocument12 paginiBryophytesAiman KaleemÎncă nu există evaluări
- Stems Lesson 1: Stem FunctionsDocument11 paginiStems Lesson 1: Stem Functionskesyah rubidoÎncă nu există evaluări
- PERIDA EMMALYN T. - BSci103-Lesson 4.2-Learning TaskDocument5 paginiPERIDA EMMALYN T. - BSci103-Lesson 4.2-Learning TaskEmmalyn PeridaÎncă nu există evaluări
- Medico Legal Aspect of DeathDocument32 paginiMedico Legal Aspect of DeathKU RU RUÎncă nu există evaluări
- Aman Human BehaviourDocument21 paginiAman Human BehaviourGurmeet BainsÎncă nu există evaluări
- RRL LocalinternationalDocument11 paginiRRL LocalinternationalkdfhjfhfÎncă nu există evaluări
- Acute MI DMII Medical ManagementDocument52 paginiAcute MI DMII Medical ManagementsherwincruzÎncă nu există evaluări
- Case Study 416Document34 paginiCase Study 416jennifer_crumm100% (1)
- 2nd Long Exam BlueprintDocument3 pagini2nd Long Exam BlueprintRachelle MaravillaÎncă nu există evaluări
- Neonatal Pneumonia Case StudyDocument9 paginiNeonatal Pneumonia Case StudyMary hope DomalaonÎncă nu există evaluări
- Log RegDocument32 paginiLog RegSIDDHARTH KUMARÎncă nu există evaluări
- 9 Dissacharides Metabolism Compatibility ModeDocument16 pagini9 Dissacharides Metabolism Compatibility ModeAubrey Nativity Ostulano YangzonÎncă nu există evaluări
- Bio Chapter 14 NotesDocument3 paginiBio Chapter 14 NotesAvehiÎncă nu există evaluări
- Chaturvedi 2004Document2 paginiChaturvedi 2004Umer HussainÎncă nu există evaluări
- Clinico-Microbiological Spectrum of Hemodialysis Catheter-Related Blood Stream Infections: A Concise ReviewDocument7 paginiClinico-Microbiological Spectrum of Hemodialysis Catheter-Related Blood Stream Infections: A Concise ReviewIJAR JOURNALÎncă nu există evaluări
- 10 12 15hematoo Edit 1Document10 pagini10 12 15hematoo Edit 1Anonymous 18GsyXbÎncă nu există evaluări
- Your Body Believes Every Word Y - Barbara Hoberman LevineDocument380 paginiYour Body Believes Every Word Y - Barbara Hoberman LevineRoryBradshawÎncă nu există evaluări
- A2 - Practice 29Document2 paginiA2 - Practice 29123VinhcuuÎncă nu există evaluări
- Study Guide STH Edition: Clinical LaboratoryDocument30 paginiStudy Guide STH Edition: Clinical LaboratoryMendoza, Richelle A.Încă nu există evaluări
- Performance Evaluation of Three Rapid Screening Assays For Detection of Antibodies To Hepatitis C Virus in CameroonDocument4 paginiPerformance Evaluation of Three Rapid Screening Assays For Detection of Antibodies To Hepatitis C Virus in CameroonVikas NakraniÎncă nu există evaluări
- March Case StudyDocument13 paginiMarch Case Studyapi-212894050Încă nu există evaluări
- Ashtanga Yoga I and II Complete OCR (Finereader - IsTA SettingsDocument192 paginiAshtanga Yoga I and II Complete OCR (Finereader - IsTA SettingsAndreea Bertea0% (1)
- A Large Prospective Open-Label Multicent PDFDocument6 paginiA Large Prospective Open-Label Multicent PDFsyeda tahsin naharÎncă nu există evaluări
- Msds ChosunDocument6 paginiMsds ChosunDheeraj E MohananÎncă nu există evaluări
- All About Lemongrass OilDocument9 paginiAll About Lemongrass OilseptiaÎncă nu există evaluări
- Copd Action Plan PDFDocument2 paginiCopd Action Plan PDFdtech2Încă nu există evaluări
- Inheritance and Selective Breeding QsDocument27 paginiInheritance and Selective Breeding QspartinpoutsamouÎncă nu există evaluări
- Molecular Biology R - F - Weaver 5th EdDocument1 paginăMolecular Biology R - F - Weaver 5th EdAlapan NandaÎncă nu există evaluări
- Tromso. Norway Orders Drug-Free Treatment in PsychiatryDocument21 paginiTromso. Norway Orders Drug-Free Treatment in PsychiatryMarcelo Astorga VelosoÎncă nu există evaluări
- Case Study Discussion (A)Document3 paginiCase Study Discussion (A)Bq dwi EvaÎncă nu există evaluări
- KetamineDocument23 paginiKetamineDimas Gatra DiantoroÎncă nu există evaluări
- TST Prep - 100 TOEFL Reading Practice Questions-110-120Document11 paginiTST Prep - 100 TOEFL Reading Practice Questions-110-120S EÎncă nu există evaluări
- Tales from Both Sides of the Brain: A Life in NeuroscienceDe la EverandTales from Both Sides of the Brain: A Life in NeuroscienceEvaluare: 3 din 5 stele3/5 (18)
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (5)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe la EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesEvaluare: 4.5 din 5 stele4.5/5 (397)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe la Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessEvaluare: 4 din 5 stele4/5 (33)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe la EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeÎncă nu există evaluări
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe la EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceEvaluare: 4.5 din 5 stele4.5/5 (517)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseDe la EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseEvaluare: 4.5 din 5 stele4.5/5 (52)
- Human: The Science Behind What Makes Your Brain UniqueDe la EverandHuman: The Science Behind What Makes Your Brain UniqueEvaluare: 3.5 din 5 stele3.5/5 (38)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDe la EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomEvaluare: 4 din 5 stele4/5 (216)
- Who's in Charge?: Free Will and the Science of the BrainDe la EverandWho's in Charge?: Free Will and the Science of the BrainEvaluare: 4 din 5 stele4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)De la EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Evaluare: 4 din 5 stele4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe la EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedEvaluare: 4 din 5 stele4/5 (11)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (812)
- Good Without God: What a Billion Nonreligious People Do BelieveDe la EverandGood Without God: What a Billion Nonreligious People Do BelieveEvaluare: 4 din 5 stele4/5 (66)
- Seven and a Half Lessons About the BrainDe la EverandSeven and a Half Lessons About the BrainEvaluare: 4 din 5 stele4/5 (110)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe la EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorÎncă nu există evaluări
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDe la EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperEvaluare: 4.5 din 5 stele4.5/5 (15)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDe la EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemEvaluare: 4.5 din 5 stele4.5/5 (115)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDe la EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouEvaluare: 4.5 din 5 stele4.5/5 (62)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignDe la EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignEvaluare: 4.5 din 5 stele4.5/5 (39)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDe la EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsEvaluare: 4.5 din 5 stele4.5/5 (2083)
- The Invention of Tomorrow: A Natural History of ForesightDe la EverandThe Invention of Tomorrow: A Natural History of ForesightEvaluare: 4.5 din 5 stele4.5/5 (5)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDe la EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldEvaluare: 4 din 5 stele4/5 (595)