S-ar putea să vă placă și
- Autoclave ExpansionDocument2 paginiAutoclave ExpansionRiscia Rapio NaduraÎncă nu există evaluări
- 2 - Straight Line DepreciationDocument1 pagină2 - Straight Line DepreciationRiscia Rapio NaduraÎncă nu există evaluări
- Philippines: Why Philippine Cinema No Longer Make As Much Action Films Like It Used To. Retrieved From Pinoy Movie BloggerDocument1 paginăPhilippines: Why Philippine Cinema No Longer Make As Much Action Films Like It Used To. Retrieved From Pinoy Movie BloggerRiscia Rapio NaduraÎncă nu există evaluări
- Airlift Vs GeyserDocument1 paginăAirlift Vs GeyserRiscia Rapio NaduraÎncă nu există evaluări
- 3dconnexion: Coordinate System (WCS)Document48 pagini3dconnexion: Coordinate System (WCS)Riscia Rapio NaduraÎncă nu există evaluări
- Second Grade Dolch WordsDocument1 paginăSecond Grade Dolch WordsRiscia Rapio NaduraÎncă nu există evaluări
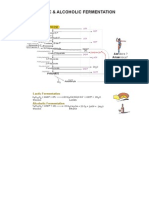
- NADURA - ChE 190 Technical Review - BoysenDocument4 paginiNADURA - ChE 190 Technical Review - BoysenRiscia Rapio NaduraÎncă nu există evaluări
- Big Yellow Taxi Salin-AwitDocument3 paginiBig Yellow Taxi Salin-AwitRiscia Rapio NaduraÎncă nu există evaluări
- Big Yellow Taxi Salin-AwitDocument3 paginiBig Yellow Taxi Salin-AwitRiscia Rapio NaduraÎncă nu există evaluări
- Mathematics ExercisesDocument2 paginiMathematics ExercisesRiscia Rapio NaduraÎncă nu există evaluări
- Samsung and South KoreaDocument2 paginiSamsung and South KoreaRiscia Rapio NaduraÎncă nu există evaluări
- SHMDocument2 paginiSHMRiscia Rapio NaduraÎncă nu există evaluări
- Nomenclature of AlkanesDocument1 paginăNomenclature of AlkanesRiscia Rapio NaduraÎncă nu există evaluări
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Recovery of Organic Acids From Fermentation BrothsDocument20 paginiRecovery of Organic Acids From Fermentation BrothsLCteyÎncă nu există evaluări
- Sick and TiredDocument24 paginiSick and TiredVaradrajan MudliyarÎncă nu există evaluări
- Unit 11 Pickles, Chutneys, Sauces and Tomato Products: StructureDocument24 paginiUnit 11 Pickles, Chutneys, Sauces and Tomato Products: StructureS NanzÎncă nu există evaluări
- Developing Fermentation Media for Industrial ProcessesDocument13 paginiDeveloping Fermentation Media for Industrial ProcessesChandiniÎncă nu există evaluări
- Aerobic Respiration WorksheetDocument7 paginiAerobic Respiration WorksheeterikabeltranÎncă nu există evaluări
- Bio 201 Laboratory ReportDocument5 paginiBio 201 Laboratory Reportapi-252855115Încă nu există evaluări
- FPJDocument1 paginăFPJThea Maria RedondoÎncă nu există evaluări
- Engineering Metabolism Through Dynamic Control: SciencedirectDocument11 paginiEngineering Metabolism Through Dynamic Control: SciencedirectShampa SenÎncă nu există evaluări
- Bioethanol Chap 1 and 2Document14 paginiBioethanol Chap 1 and 2Rubelynn Marthe0% (1)
- Alex TellaDocument24 paginiAlex TellaDesta Tesfay100% (2)
- Termossonicação Nondairy ResultadosDocument9 paginiTermossonicação Nondairy ResultadosBruna VieiraÎncă nu există evaluări
- Best Compost Tea Guide Under 40 CharactersDocument34 paginiBest Compost Tea Guide Under 40 CharactersWes D Gordon100% (1)
- 10 1016@j Foodres 2021 110482Document20 pagini10 1016@j Foodres 2021 110482Daniel ParedesÎncă nu există evaluări
- Food MicrobiologyDocument63 paginiFood MicrobiologyFaye EbuenÎncă nu există evaluări
- Lesson 10: Sterilization: Learning ObjectivesDocument8 paginiLesson 10: Sterilization: Learning Objectivesrohit rajputÎncă nu există evaluări
- Respiration AristoDocument58 paginiRespiration AristoMohamed Attia Imam SafanÎncă nu există evaluări
- Natural and Cultured Buttermilk: Ravinder Kumar Anita Kumari GarsaDocument25 paginiNatural and Cultured Buttermilk: Ravinder Kumar Anita Kumari GarsaUyên SandyÎncă nu există evaluări
- Rice Beer Biochemical AnalysisDocument7 paginiRice Beer Biochemical AnalysisIrati PalÎncă nu există evaluări
- Application of Indigenous Microorganisms (Imos) On Poultry FloorDocument6 paginiApplication of Indigenous Microorganisms (Imos) On Poultry FloorSriram BhupaathirajuÎncă nu există evaluări
- Lactic and Alcoholic FermentationDocument1 paginăLactic and Alcoholic FermentationDr. SHIVA AITHALÎncă nu există evaluări
- LAS - BIOTECH Q2 WK 1-2Document13 paginiLAS - BIOTECH Q2 WK 1-2Febie Palapuz-AgbayaniÎncă nu există evaluări
- Thosai FermentationDocument15 paginiThosai FermentationVithyia Murugiah100% (1)
- Lesson 2 Aerobic Vs Anaerobic Respiration-1Document12 paginiLesson 2 Aerobic Vs Anaerobic Respiration-1John denver Arenas100% (1)
- SawyerrOkudoh2019 IntJEnergyEconPolicyDocument13 paginiSawyerrOkudoh2019 IntJEnergyEconPolicyHj JayatheerthaÎncă nu există evaluări
- Growth of Probiotic in Soy Yogurt Formulation PDFDocument8 paginiGrowth of Probiotic in Soy Yogurt Formulation PDFLenaldy Nuari GarnokoÎncă nu există evaluări
- Food Microbiology: The Basics and The Details of Cheese ProductionDocument76 paginiFood Microbiology: The Basics and The Details of Cheese ProductionbhaleshÎncă nu există evaluări
- Fermentation Lesson Plan 1Document3 paginiFermentation Lesson Plan 1api-571580523Încă nu există evaluări
- The Bacteria, 1880Document260 paginiThe Bacteria, 1880deryhermawanÎncă nu există evaluări
- CELLULAR ENERGYDocument2 paginiCELLULAR ENERGYJohn Daniel AntolinÎncă nu există evaluări
- Alcohols & Carboxylic Acids 1 QPDocument12 paginiAlcohols & Carboxylic Acids 1 QPShiv DholuÎncă nu există evaluări