S-ar putea să vă placă și
- Corporate Brochure Ar PDFDocument16 paginiCorporate Brochure Ar PDFReham IbrahimÎncă nu există evaluări
- Genetic diversity of medicinal plants using molecular markersDocument12 paginiGenetic diversity of medicinal plants using molecular markersReham IbrahimÎncă nu există evaluări
- Halmouch 8 3 2015 232 237 PDFDocument6 paginiHalmouch 8 3 2015 232 237 PDFReham IbrahimÎncă nu există evaluări
- Genetic diversity of medicinal plants using molecular markersDocument12 paginiGenetic diversity of medicinal plants using molecular markersReham IbrahimÎncă nu există evaluări
- Agro 2013Document31 paginiAgro 2013Reham IbrahimÎncă nu există evaluări
- Genetic Engineering of Plants via AgrobacteriumDocument21 paginiGenetic Engineering of Plants via AgrobacteriumReham IbrahimÎncă nu există evaluări
- Agrobacterium Mediated Transformation of Sompositae Plants I Construction of Transgenic Plants and Hairy Roots With New Properties PDFDocument13 paginiAgrobacterium Mediated Transformation of Sompositae Plants I Construction of Transgenic Plants and Hairy Roots With New Properties PDFReham IbrahimÎncă nu există evaluări
- Epigenetics PDFDocument50 paginiEpigenetics PDFReham Ibrahim75% (4)
- Evolution & Speciation PDFDocument4 paginiEvolution & Speciation PDFReham IbrahimÎncă nu există evaluări
- Genetic ImproveDocument83 paginiGenetic ImproveReham IbrahimÎncă nu există evaluări
- Tar 1406 144 PDFDocument12 paginiTar 1406 144 PDFReham IbrahimÎncă nu există evaluări
- Applications of Genetics To Conservation Biology Molecular TaxonomyDocument10 paginiApplications of Genetics To Conservation Biology Molecular TaxonomyReham IbrahimÎncă nu există evaluări
- Crop Improvement PDFDocument2 paginiCrop Improvement PDFReham IbrahimÎncă nu există evaluări
- Diversity: Assessing Plant Genetic Diversity by Molecular ToolsDocument17 paginiDiversity: Assessing Plant Genetic Diversity by Molecular ToolsErlyta ViviÎncă nu există evaluări
- A Training Module For The International Course On The Management and Utilisation of Field Genebanks and in Vitro Collections 1643 PDFDocument100 paginiA Training Module For The International Course On The Management and Utilisation of Field Genebanks and in Vitro Collections 1643 PDFReham IbrahimÎncă nu există evaluări
- PresentationDocument75 paginiPresentationReham IbrahimÎncă nu există evaluări
- A Training Module For The International Course On Plant Genetic Resources and Genebank Management 1644 PDFDocument88 paginiA Training Module For The International Course On Plant Genetic Resources and Genebank Management 1644 PDFReham IbrahimÎncă nu există evaluări
- Sequencing Genes and GenomesDocument39 paginiSequencing Genes and GenomesReham IbrahimÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Botany (Flower)Document15 paginiBotany (Flower)Louis Fetilo FabunanÎncă nu există evaluări
- Bio CH6 F5 StudywithadminDocument7 paginiBio CH6 F5 StudywithadminRathi MalarÎncă nu există evaluări
- Pollen-Pistil Interaction: Pollen Attachment and HydrationDocument6 paginiPollen-Pistil Interaction: Pollen Attachment and HydrationSreeja Raj100% (1)
- Science Hints & Solutions: Series JBB/4Document11 paginiScience Hints & Solutions: Series JBB/4Paplu MasterÎncă nu există evaluări
- CBSE Biology Question Bank on Sexual Reproduction in Flowering PlantsDocument12 paginiCBSE Biology Question Bank on Sexual Reproduction in Flowering PlantsMohamed DhaanishÎncă nu există evaluări
- BC 2017 Lecture 2 Dixit Plant Cell BiologyDocument29 paginiBC 2017 Lecture 2 Dixit Plant Cell BiologyAishwarya MondalÎncă nu există evaluări
- Bergamo Et Al. 2020 - Negative Density-DependenceDocument11 paginiBergamo Et Al. 2020 - Negative Density-DependenceGudryan J. BaronioÎncă nu există evaluări
- Class12 Bio QB Part 1Document83 paginiClass12 Bio QB Part 1Rohit KumarÎncă nu există evaluări
- Anther Dehiscence, Pollen Viability and Stigma Receptivity Study On Cultivars of Black Pepper (Piper Nigrum L.)Document14 paginiAnther Dehiscence, Pollen Viability and Stigma Receptivity Study On Cultivars of Black Pepper (Piper Nigrum L.)phung nguyen100% (1)
- Mechanical Design of Pollen AperturesDocument4 paginiMechanical Design of Pollen AperturesAntonio ŠiberÎncă nu există evaluări
- Biology Investigatory ProjectDocument19 paginiBiology Investigatory ProjectPriyanks Rout50% (2)
- The Learners Should Be Able To Create A Hypothetical Community To Show How Organisms Interact and Reproduce To SurviveDocument9 paginiThe Learners Should Be Able To Create A Hypothetical Community To Show How Organisms Interact and Reproduce To SurviveMaria Jonalyn SilvanoÎncă nu există evaluări
- Reproduction in Plants: Test Yourself 17.1 (Page 316)Document3 paginiReproduction in Plants: Test Yourself 17.1 (Page 316)lee100% (1)
- Meiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inDocument29 paginiMeiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inyamamaÎncă nu există evaluări
- Pollination Types and ProcessDocument22 paginiPollination Types and ProcessNAVEEN DHIMAN100% (1)
- SPM Biology Essays CollectionDocument31 paginiSPM Biology Essays CollectionYvonne Choo Shuen Lann100% (49)
- Bio ProjectDocument13 paginiBio ProjectFeLiX JeremyÎncă nu există evaluări
- Sexual IncompatibilityDocument3 paginiSexual IncompatibilitySanjoy NingthoujamÎncă nu există evaluări
- Chapter - 12 Reproduction in PlantsDocument5 paginiChapter - 12 Reproduction in Plantsanju singhÎncă nu există evaluări
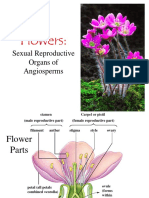
- Flowers: Sexual Reproduction in AngiospermsDocument106 paginiFlowers: Sexual Reproduction in AngiospermsAlfonso PlantillaÎncă nu există evaluări
- Sita 2017Document30 paginiSita 2017skpuindia7Încă nu există evaluări
- Seed Saving GuideDocument30 paginiSeed Saving GuideAntanas Brudinys100% (5)
- 13 Methods To Overcome Self-Incompatibility in Plants: Method # 1. Bud PollinationDocument9 pagini13 Methods To Overcome Self-Incompatibility in Plants: Method # 1. Bud PollinationManshi GhansalaÎncă nu există evaluări
- Programmed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciDocument23 paginiProgrammed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciSourav GuptaÎncă nu există evaluări
- Pollen Viability and in Vitro Pollen Germination Studies in Momordica Species and Their Intra and Interspecific HybridsDocument10 paginiPollen Viability and in Vitro Pollen Germination Studies in Momordica Species and Their Intra and Interspecific HybridsJagatha RaasmiÎncă nu există evaluări
- Answers: Plant Reproduction - Pollination and Fertilisation Teaching NotesDocument5 paginiAnswers: Plant Reproduction - Pollination and Fertilisation Teaching NotesKhoo Ying WeiÎncă nu există evaluări
- Class 12 NEET Revision Notes - 7083332 PDFDocument146 paginiClass 12 NEET Revision Notes - 7083332 PDFvengateshwaran kÎncă nu există evaluări
- Pollination and Fertilisation Question BankDocument17 paginiPollination and Fertilisation Question BankRonnith Nandy100% (1)
- Gnetum GnemonDocument15 paginiGnetum GnemonmastikaÎncă nu există evaluări
- Biology Project Class 12Document17 paginiBiology Project Class 12RakeshÎncă nu există evaluări