S-ar putea să vă placă și
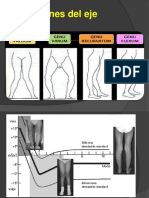
- Genu VaroDocument8 paginiGenu VaroArsenio Flores VasquezÎncă nu există evaluări
- Manual de Neumologia ClinicaDocument337 paginiManual de Neumologia ClinicaRicardo Casillas Flores100% (4)
- 04 Ulceras CornealesDocument25 pagini04 Ulceras CornealesArsenio Flores VasquezÎncă nu există evaluări
- 04 Ulceras Corneales PDFDocument3 pagini04 Ulceras Corneales PDFArsenio Flores VasquezÎncă nu există evaluări
- Historia Clinica Orientada A ProblemasDocument14 paginiHistoria Clinica Orientada A ProblemasFranz HuamaniÎncă nu există evaluări
- Expo Tecnica SeminarioDocument19 paginiExpo Tecnica SeminarioArsenio Flores VasquezÎncă nu există evaluări
- Notas 2Document1 paginăNotas 2Arsenio Flores VasquezÎncă nu există evaluări
- TVM HomicDocument67 paginiTVM HomicArsenio Flores VasquezÎncă nu există evaluări
- Sillabus de Cirugía IIDocument33 paginiSillabus de Cirugía IIArsenio Flores VasquezÎncă nu există evaluări
- ValeDocument13 paginiValealexÎncă nu există evaluări
- Patologia-Anorectal-benigna - NNN, Napoleón GonzalesDocument62 paginiPatologia-Anorectal-benigna - NNN, Napoleón GonzalesArsenio Flores VasquezÎncă nu există evaluări
- Toma de Muestra de OrinaDocument70 paginiToma de Muestra de OrinamireÎncă nu există evaluări
- Parpados y Conjuntiva.Document30 paginiParpados y Conjuntiva.Arsenio Flores Vasquez100% (1)
- Aminoglucósidos VDocument3 paginiAminoglucósidos VArsenio Flores VasquezÎncă nu există evaluări
- 1 Osteoartritis30sept2011 111125033933 Phpapp01Document61 pagini1 Osteoartritis30sept2011 111125033933 Phpapp01Arsenio Flores VasquezÎncă nu există evaluări
- SUSTENTACIÓNDocument1 paginăSUSTENTACIÓNArsenio Flores VasquezÎncă nu există evaluări
- Terminología MédicaDocument25 paginiTerminología MédicaLalo Canalizo Veliz67% (3)
- Gases Arteriales - Joe.2012Document42 paginiGases Arteriales - Joe.2012Alexander Campos TapiaÎncă nu există evaluări
- Administracion Recursos HumanosDocument7 paginiAdministracion Recursos HumanosArsenio Flores VasquezÎncă nu există evaluări
- GERIATRÍA-Fundamentos de La GeriatríaDocument55 paginiGERIATRÍA-Fundamentos de La GeriatríaArsenio Flores Vasquez0% (1)
- Formas ClinicasDocument25 paginiFormas ClinicasArsenio Flores VasquezÎncă nu există evaluări
- RESPIRATORIODocument30 paginiRESPIRATORIOArsenio Flores VasquezÎncă nu există evaluări
- Radiología torácica normalDocument56 paginiRadiología torácica normalVictor RiquelmeÎncă nu există evaluări
- GuacalaDocument2 paginiGuacalaArsenio Flores VasquezÎncă nu există evaluări
- HCCP Graciela Esquen ChavezDocument10 paginiHCCP Graciela Esquen ChavezArsenio Flores VasquezÎncă nu există evaluări
- PancitopeniaDocument17 paginiPancitopeniaBenjamin Torres Octavo100% (1)
- TBCDocument74 paginiTBCArsenio Flores VasquezÎncă nu există evaluări
- ABC Diagnostico Diferencial de Emergencia PDFDocument102 paginiABC Diagnostico Diferencial de Emergencia PDFLina Marcela PortillaÎncă nu există evaluări
- Morfología Del Conducto Torácico y Su Importancia Clínica: Artículo de RevisiónDocument5 paginiMorfología Del Conducto Torácico y Su Importancia Clínica: Artículo de RevisiónHenry Daniel MarigordaÎncă nu există evaluări
- Test Cirugia Toracica y NeumoDocument23 paginiTest Cirugia Toracica y NeumoLucio RubiÎncă nu există evaluări
- Producción de Metanol A Base de MetanoDocument7 paginiProducción de Metanol A Base de MetanoValentina Angelica Concha RamirezÎncă nu există evaluări
- Anexo 1 - Tarea 3 - Enzimología y BioenergéticaDocument10 paginiAnexo 1 - Tarea 3 - Enzimología y BioenergéticaLeidy Johana BravoÎncă nu există evaluări
- Genética prácticaDocument8 paginiGenética prácticaManuel Alonzo Eizaguirre MontesÎncă nu există evaluări
- Catalogo Toricas Quadring Es fr3Document40 paginiCatalogo Toricas Quadring Es fr3alejandro obregonÎncă nu există evaluări
- Practica 6. Desnaturalización de Proteinas - Guía para El ProfesorDocument6 paginiPractica 6. Desnaturalización de Proteinas - Guía para El ProfesorDannyIsaacPaucarimaÎncă nu există evaluări
- Documento Base Glosario Biología 2º BACHDocument6 paginiDocumento Base Glosario Biología 2º BACHPaula Padilla BermúdezÎncă nu există evaluări
- Colageno ProyectooDocument27 paginiColageno ProyectooAlecitaGutierrezMartinez100% (1)
- AzlactonaDocument27 paginiAzlactonaJulissa Minaya Aparicio100% (1)
- Cellphone Cover and HipersensitivityDocument2 paginiCellphone Cover and HipersensitivityPayGamer YTÎncă nu există evaluări
- Balance de Masa para Bioprocesos Con Crecimiento Celular TeoríaDocument34 paginiBalance de Masa para Bioprocesos Con Crecimiento Celular TeoríaRAMOS OLLACHICA MARYORI JENNIFERÎncă nu există evaluări
- QuizzDocument5 paginiQuizzFrederick HeisenbergÎncă nu există evaluări
- Acidos Nucleicos ResumenDocument16 paginiAcidos Nucleicos ResumenFabio VilladiegoÎncă nu există evaluări
- Ficha Tecnica Alcohol 96° Bioxclean - OficialDocument10 paginiFicha Tecnica Alcohol 96° Bioxclean - OficialSaymon Bautista Tapia100% (1)
- ISOMEROSDocument10 paginiISOMEROSGabriela PradoÎncă nu există evaluări
- Aromaticidad PDFDocument12 paginiAromaticidad PDFjuan manuel rodriguezÎncă nu există evaluări
- AuxinasDocument12 paginiAuxinaslalalala123Încă nu există evaluări
- Citogenetica 3 WDocument78 paginiCitogenetica 3 WadrianÎncă nu există evaluări
- Clase 5. Fosforilación OxidativaDocument44 paginiClase 5. Fosforilación OxidativaRafael Mendoza RodriguezÎncă nu există evaluări
- Los Aldehídos Son Compuestos Químicos de Tipo Orgánico Que Surgen Cuando Determinados Alcoholes Se OxidanDocument2 paginiLos Aldehídos Son Compuestos Químicos de Tipo Orgánico Que Surgen Cuando Determinados Alcoholes Se OxidanVicky LopezÎncă nu există evaluări
- Reglas de Nomenclatura Iupac para Escribir Nombres y Formulas de Los Compuestos Orgánicos Oxigenados y Halogenuros de AlquiloDocument17 paginiReglas de Nomenclatura Iupac para Escribir Nombres y Formulas de Los Compuestos Orgánicos Oxigenados y Halogenuros de AlquiloHJMPPÎncă nu există evaluări
- Propiedades de Especies PurasDocument66 paginiPropiedades de Especies PurasBrian WilliamsÎncă nu există evaluări
- Correccion de Practica 1 Sintesis de BenzopinacolDocument11 paginiCorreccion de Practica 1 Sintesis de BenzopinacolsandramaldonadoÎncă nu există evaluări
- Aceite Esencial AnetolDocument3 paginiAceite Esencial Anetolanon_801554080Încă nu există evaluări
- Síntesis de 2,3-difenilquinoxalina mediante química verdeDocument7 paginiSíntesis de 2,3-difenilquinoxalina mediante química verdeIvanÎncă nu există evaluări
- Polimeros 1Document7 paginiPolimeros 1Josue Antonio Torres CentenoÎncă nu există evaluări
- Chavez GuzmanCristalizacionDocument5 paginiChavez GuzmanCristalizacionCami ChavezÎncă nu există evaluări
- PolicarbonatoDocument8 paginiPolicarbonatoJessica Rmz NietoÎncă nu există evaluări
- Estructura y Nomenclatura de Los ÉteresDocument9 paginiEstructura y Nomenclatura de Los ÉteresLuis GarciaÎncă nu există evaluări
- 1726 Cuestionario PolimerosDocument3 pagini1726 Cuestionario PolimerosClaydÎncă nu există evaluări
- Electroforesis de ADN en Geles de AgarosaDocument4 paginiElectroforesis de ADN en Geles de AgarosaCaro RamírezÎncă nu există evaluări