S-ar putea să vă placă și
- Borboletas E Mariposas De Santa CatarinaDe la EverandBorboletas E Mariposas De Santa CatarinaÎncă nu există evaluări
- Nanopartículas Magnéticas: Um Estudo EstruturalDe la EverandNanopartículas Magnéticas: Um Estudo EstruturalÎncă nu există evaluări
- Apostila Amphibia PDFDocument57 paginiApostila Amphibia PDFRonaldo Leite Da Silva Filho LeiteÎncă nu există evaluări
- ZoologiaDocument105 paginiZoologiawanessas_sol100% (3)
- Zoologia de VertebradosDocument225 paginiZoologia de VertebradosDaniel RochaÎncă nu există evaluări
- Proposta para Aulas de Invertebrados Ef PDFDocument53 paginiProposta para Aulas de Invertebrados Ef PDFAndreza MartinsÎncă nu există evaluări
- Raios Catodicos MioloDocument96 paginiRaios Catodicos MioloMarta CharnecaÎncă nu există evaluări
- Sobre A Laterita - POTENCIALIDADE DO CONGLOMERADO FERRUGINOSO LATERÍTICODocument344 paginiSobre A Laterita - POTENCIALIDADE DO CONGLOMERADO FERRUGINOSO LATERÍTICOLucas RamisÎncă nu există evaluări
- Morfologia Da Língua Portuguesa Vol 1 - Marcelo Moraes CaetanoDocument93 paginiMorfologia Da Língua Portuguesa Vol 1 - Marcelo Moraes CaetanoAcostaÎncă nu există evaluări
- A APLICAÇÃO DA PALEOARTE E DO DESENHO CIENTÍFICO NA REPRESENTAÇÃO DA CLASSE REPTILIA Victor Feijó de CarvalhoDocument114 paginiA APLICAÇÃO DA PALEOARTE E DO DESENHO CIENTÍFICO NA REPRESENTAÇÃO DA CLASSE REPTILIA Victor Feijó de CarvalhoVictor FeijóÎncă nu există evaluări
- Relatório de Práticas em Zoologia de VertebradosDocument8 paginiRelatório de Práticas em Zoologia de VertebradosJoão Pedro Alves GoulartÎncă nu există evaluări
- Agente e Escrivao Da Policia Federal Aula 1 Arquivologia Sormany AlvesDocument14 paginiAgente e Escrivao Da Policia Federal Aula 1 Arquivologia Sormany AlvesRafael SantanaÎncă nu există evaluări
- Cultivo de VieiraDocument156 paginiCultivo de VieiraClárence Gibson100% (1)
- Invmar 1Document87 paginiInvmar 1Daiane Armstrong FukuyiÎncă nu există evaluări
- Ulfl254195 TMDocument132 paginiUlfl254195 TMNelsonynelsonÎncă nu există evaluări
- Manual de Linguistica 1 PDFDocument250 paginiManual de Linguistica 1 PDFKarlla Ribeiro100% (1)
- Apostila AgrometeorologiaDocument153 paginiApostila AgrometeorologiaRaquel Junqueira De CastroÎncă nu există evaluări
- Informe Sobre As Espècies Exûticas Marinhas No Brasil PDFDocument440 paginiInforme Sobre As Espècies Exûticas Marinhas No Brasil PDFNatália GomesÎncă nu există evaluări
- Tecnologia PesqueiraDocument54 paginiTecnologia Pesqueiraangela ferreiraÎncă nu există evaluări
- Gambiarra e Experimentalismo Sonoro, Giuliano Obici. 2014Document194 paginiGambiarra e Experimentalismo Sonoro, Giuliano Obici. 2014Gabriela FavreÎncă nu există evaluări
- BASSETTO (2005, v.1)Document383 paginiBASSETTO (2005, v.1)Kauã MatassoliÎncă nu există evaluări
- História Da Técnica e Da Tecnologia - Textos Básicos by Ruy GamaDocument278 paginiHistória Da Técnica e Da Tecnologia - Textos Básicos by Ruy GamaKelly MedinaÎncă nu există evaluări
- Ensino e Aprendizagem de Português No Mercosul PDFDocument211 paginiEnsino e Aprendizagem de Português No Mercosul PDFCarolina ContrerasÎncă nu există evaluări
- E Sample 97885352880186Document20 paginiE Sample 97885352880186nsanieleÎncă nu există evaluări
- Catalogo Das Especies de Peixes MarinhosDocument164 paginiCatalogo Das Especies de Peixes Marinhosdia100noiteÎncă nu există evaluări
- Introdução A Taxonomia de FormigasDocument15 paginiIntrodução A Taxonomia de FormigasVALTERJRAJUÎncă nu există evaluări
- Relatorio Zoo II Excursao Xai XaiDocument14 paginiRelatorio Zoo II Excursao Xai XaiAlson Carlos100% (1)
- Fundo RIDocument58 paginiFundo RIjessicamaximo6Încă nu există evaluări
- Tecnologia Do PesacdoDocument212 paginiTecnologia Do PesacdoMiguel Teixeira Couteiro100% (1)
- Relatorio PerifitonDocument18 paginiRelatorio PerifitonJulio CesarÎncă nu există evaluări
- Pedologia AplicadaDocument147 paginiPedologia AplicadaEDIVALDO LISBOA AGROÎncă nu există evaluări
- Historia Do PortuguêsDocument8 paginiHistoria Do Portuguêsfb.tnr100% (1)
- Ditongos Orais No PeDocument124 paginiDitongos Orais No PealinelatinaÎncă nu există evaluări
- Revisão Do Gênero Tetanorhynchus Brunner Von Wattenwyl, 1890 OrthopteraDocument204 paginiRevisão Do Gênero Tetanorhynchus Brunner Von Wattenwyl, 1890 OrthopteraFgtRafaelFernandesÎncă nu există evaluări
- Comportamentos de Uma Populacao de Pato-Real Residente Num Parque Urbano No Norte de Portugal Du2Document83 paginiComportamentos de Uma Populacao de Pato-Real Residente Num Parque Urbano No Norte de Portugal Du2GUHPYÎncă nu există evaluări
- 2006 - Chave BicudoDocument4 pagini2006 - Chave BicudoIsabel MonteiroÎncă nu există evaluări
- Tese - Algoritmos de Processamento de Linguagem Natural Na Síntese Da Fala - Daniela Filipa Macedo Moreira Da Silva Braga (2008)Document228 paginiTese - Algoritmos de Processamento de Linguagem Natural Na Síntese Da Fala - Daniela Filipa Macedo Moreira Da Silva Braga (2008)Llama VicuñaÎncă nu există evaluări
- Morcegos em Áreas Urbanas e Rurais - Manual de Manejo e ControleDocument116 paginiMorcegos em Áreas Urbanas e Rurais - Manual de Manejo e ControleMarcos Vinícius93% (28)
- Instructivo Practica 5 Reino Animal Vertebrados b2b Efpem 2015Document6 paginiInstructivo Practica 5 Reino Animal Vertebrados b2b Efpem 2015Anonymous SfLVmdÎncă nu există evaluări
- Nova Gramatica Do Portugues Brasileiro SumarioDocument17 paginiNova Gramatica Do Portugues Brasileiro SumarioVinicius Neves100% (1)
- Suzana Ii Trabalho de Zoologia Geral Editado PDFDocument15 paginiSuzana Ii Trabalho de Zoologia Geral Editado PDFpaulo M. JulioÎncă nu există evaluări
- Relatorio Biologia AquaticaDocument10 paginiRelatorio Biologia AquaticaLivia Costa de SousaÎncă nu există evaluări
- A Classificação e A Avaliação Documental Na MarinhaDocument93 paginiA Classificação e A Avaliação Documental Na MarinhaSérgio SimõesÎncă nu există evaluări
- PANAFTOSA 2007 Manual de Procedimentos para A Atencao As Ocorrencias de Febre Aftosa e Outras Enfermidades VesicularesDocument145 paginiPANAFTOSA 2007 Manual de Procedimentos para A Atencao As Ocorrencias de Febre Aftosa e Outras Enfermidades VesicularesigorvetÎncă nu există evaluări
- Medicina de MamiferosDocument151 paginiMedicina de MamiferosDr.Dário Divino Dias100% (1)
- Padrões de Dedos, Uma Contribuição À Técnica Violinistica PDFDocument158 paginiPadrões de Dedos, Uma Contribuição À Técnica Violinistica PDFrntodrosa674100% (1)
- Livro+Projeto+de+Reservatorios+e+Piscinas EnsonDocument105 paginiLivro+Projeto+de+Reservatorios+e+Piscinas EnsonAnderson Ourives75% (4)
- Dicionário Multilingue de Regência Verbal PDFDocument313 paginiDicionário Multilingue de Regência Verbal PDFMarcos ViniciusÎncă nu există evaluări
- Efeito de Armadilha Das Passagens em Grelha Sobre Os AnfíbiosDocument61 paginiEfeito de Armadilha Das Passagens em Grelha Sobre Os AnfíbiosdrnitroÎncă nu există evaluări
- Práticas de Mecânica Dos SolosDocument185 paginiPráticas de Mecânica Dos SolosThiago Laurindo 2Încă nu există evaluări
- ISSUUDocument48 paginiISSUUAmanda Vieira AngeloÎncă nu există evaluări
- Guia Didatico - Formação e Caracterização Dos SolosDocument62 paginiGuia Didatico - Formação e Caracterização Dos SolosAnna SanttosÎncă nu există evaluări
- Gustavo Ribas Curcio PDFDocument510 paginiGustavo Ribas Curcio PDFEdivando Vitor Couto100% (1)
- As Políticas Linguísticas - Louis-Jean CalvetDocument167 paginiAs Políticas Linguísticas - Louis-Jean Calvetpamellar3008Încă nu există evaluări
- Apostila Completa Zoologia Dos Vertebrados FaveniDocument60 paginiApostila Completa Zoologia Dos Vertebrados Favenijany SantosÎncă nu există evaluări
- Insetos Do Oeste De Santa CatarinaDe la EverandInsetos Do Oeste De Santa CatarinaÎncă nu există evaluări
- Introdução À História Da Filosofia IDe la EverandIntrodução À História Da Filosofia IÎncă nu există evaluări
- Aula 2 - Conceito e Natureza Da Comunidade PDFDocument32 paginiAula 2 - Conceito e Natureza Da Comunidade PDFCláudia WincklerÎncă nu există evaluări
- Aula Campo 2015Document1 paginăAula Campo 2015Cláudia WincklerÎncă nu există evaluări
- Projeto PiolhoDocument11 paginiProjeto PiolhoCláudia WincklerÎncă nu există evaluări
- Piolho Prevenir Tratar PDFDocument2 paginiPiolho Prevenir Tratar PDFCláudia WincklerÎncă nu există evaluări
- 2015-2 - Guia de Estudos Ética e SociedadeDocument131 pagini2015-2 - Guia de Estudos Ética e SociedadeCláudia WincklerÎncă nu există evaluări
- 2015-2 - Guia de Estudos Ética e SociedadeDocument131 pagini2015-2 - Guia de Estudos Ética e SociedadeCláudia WincklerÎncă nu există evaluări
- 1 Ecologia Indivíduo, Ciclo de VidaDocument10 pagini1 Ecologia Indivíduo, Ciclo de VidaCláudia WincklerÎncă nu există evaluări
- Paper 3 PeriodoDocument3 paginiPaper 3 PeriodoCláudia WincklerÎncă nu există evaluări
- Paper 3 PeriodoDocument3 paginiPaper 3 PeriodoCláudia WincklerÎncă nu există evaluări
- Ameixa de InvernoDocument15 paginiAmeixa de InvernoCláudia WincklerÎncă nu există evaluări
- O Professor Como MediadorDocument5 paginiO Professor Como MediadorCláudia WincklerÎncă nu există evaluări
- LeiteDocument53 paginiLeiteJCMAntunesÎncă nu există evaluări
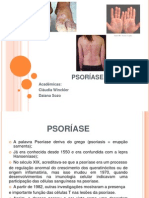
- PSORÍASEDocument19 paginiPSORÍASECláudia WincklerÎncă nu există evaluări
- Histologia 131207213055 Phpapp01Document116 paginiHistologia 131207213055 Phpapp01ClaudiaWincklerÎncă nu există evaluări
- 500 Segredos Culinários ReveladosDocument29 pagini500 Segredos Culinários ReveladosProf. Paulo Vasco100% (1)
- Distensão Muscular Entorse e LuzaçãoDocument27 paginiDistensão Muscular Entorse e Luzaçãotaati2013Încă nu există evaluări
- Tecn Controle PoluicaoDocument44 paginiTecn Controle PoluicaoCláudia WincklerÎncă nu există evaluări
- Modelagem 2Document2 paginiModelagem 2Cláudia WincklerÎncă nu există evaluări
- LeiteDocument53 paginiLeiteJCMAntunesÎncă nu există evaluări
- POP Reabilitacao Interdiscilplinar em Cuidados Prolongados e Paliativos em Neonatologia e Pediatria No Formato EBSERH 3 PDFDocument15 paginiPOP Reabilitacao Interdiscilplinar em Cuidados Prolongados e Paliativos em Neonatologia e Pediatria No Formato EBSERH 3 PDFRafael SenaÎncă nu există evaluări
- 10 Homeostasia HidroeletrolíticaDocument36 pagini10 Homeostasia HidroeletrolíticaWenderson Morais100% (1)
- Urologia Cap2 PDFDocument7 paginiUrologia Cap2 PDFLucimar FonsecaÎncă nu există evaluări
- A Imagem Do Corpo No Teatro Das Emoções e Dos Sentimentos - Sérgio GomesDocument30 paginiA Imagem Do Corpo No Teatro Das Emoções e Dos Sentimentos - Sérgio GomesErick VieiraÎncă nu există evaluări
- Saúde - TabagismoDocument20 paginiSaúde - TabagismovanisgioÎncă nu există evaluări
- ProteínasDocument3 paginiProteínasFlávio Zica100% (1)
- Sistema Digestório e Suas Respectivas FunçõesDocument29 paginiSistema Digestório e Suas Respectivas FunçõesmmmdeandradeÎncă nu există evaluări
- PRÍONSDocument10 paginiPRÍONSDani RezendeÎncă nu există evaluări
- Filo CnidariaDocument38 paginiFilo CnidariaGuilherme Rocha100% (1)
- A Fibra Muscular e Fatores Que Interferem No Seu FenótipoDocument6 paginiA Fibra Muscular e Fatores Que Interferem No Seu FenótipoSergio FalciÎncă nu există evaluări
- Fluorescência Da ClorofilaDocument14 paginiFluorescência Da ClorofilaLindomar Souza MachadoÎncă nu există evaluări
- Atividades Fisiologia - Conceitos Introdutórios Prof. Dr. Daniel Sol Sol de Medeiros HomeostasiaDocument5 paginiAtividades Fisiologia - Conceitos Introdutórios Prof. Dr. Daniel Sol Sol de Medeiros HomeostasiaWilliam JorgeÎncă nu există evaluări
- PDF DR Paulo Henrique-CoagulogramaDocument19 paginiPDF DR Paulo Henrique-CoagulogramaStella Rosa100% (1)
- O Que Ocorre Com o Organismo Durante o Jejum ProlongadoDocument1 paginăO Que Ocorre Com o Organismo Durante o Jejum Prolongadoorkut2008zuat100% (1)
- Pesquisa de Anticorpos IrregularesDocument30 paginiPesquisa de Anticorpos IrregularesJosé Menezes de Queiroz100% (1)
- ESTUDO DIRIGIDO CartilagemDocument2 paginiESTUDO DIRIGIDO CartilagemLucas CorreiaÎncă nu există evaluări
- In Olo Gias e Abre Via Çõ: + Anotações de EnfermagemDocument19 paginiIn Olo Gias e Abre Via Çõ: + Anotações de EnfermagemAnna KarolinaÎncă nu există evaluări
- Diretriz AVE IDocument13 paginiDiretriz AVE IArtur Costa BorgesÎncă nu există evaluări
- Planilha 21km Iniciante 16sDocument3 paginiPlanilha 21km Iniciante 16sMauricio MaranÎncă nu există evaluări
- Histologia Do Sistema CirculatórioDocument19 paginiHistologia Do Sistema CirculatórioMinerva AgataÎncă nu există evaluări
- Curva de Dissociação Da O2Document4 paginiCurva de Dissociação Da O2paulo100% (1)
- Teste de Recuperação 9ºDocument5 paginiTeste de Recuperação 9ºMargarida ArriagaÎncă nu există evaluări
- Impressionismo e Pos Impressionismo-1Document1 paginăImpressionismo e Pos Impressionismo-1Moisés MarquesÎncă nu există evaluări
- 2 Definiçoes de Massagens e Suas OrigensDocument32 pagini2 Definiçoes de Massagens e Suas OrigensSilvia RodriguesÎncă nu există evaluări
- Metabolismo de Carboidratos - Rosiane Gomes Silva OliveiraDocument41 paginiMetabolismo de Carboidratos - Rosiane Gomes Silva OliveiraRosiane Gomes Silva Oliveira100% (1)
- Aula 3 - Sistema OsteomuscularDocument34 paginiAula 3 - Sistema OsteomuscularFabio Delfim JuliasseÎncă nu există evaluări
- 2.PDF Pratica de FilosofiaDocument89 pagini2.PDF Pratica de FilosofiaSheila SampaioÎncă nu există evaluări
- RituaisDocument6 paginiRituaisMeloÎncă nu există evaluări
- Livro - A Força Curativa Da Respiração (Marietta Till)Document51 paginiLivro - A Força Curativa Da Respiração (Marietta Till)maguszareth100% (7)
- 011 Tratamento Da Fratura-Luxacao Trans-Escafoperisemilunar Do Carpo e Da Luxação Perisemilunar Do CarpoDocument6 pagini011 Tratamento Da Fratura-Luxacao Trans-Escafoperisemilunar Do Carpo e Da Luxação Perisemilunar Do CarpoRicardo NunesÎncă nu există evaluări