S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Burns Management PDFDocument7 paginiBurns Management PDFRoh Bungaria N Garingging100% (1)
- Anti Bullying 2013 AmmendmentDocument5 paginiAnti Bullying 2013 AmmendmentJames Matthew MadridÎncă nu există evaluări
- BambooDocument5 paginiBambooJames Matthew MadridÎncă nu există evaluări
- NSAID LectureDocument44 paginiNSAID Lecturemamun_ruÎncă nu există evaluări
- NSAID LectureDocument44 paginiNSAID Lecturemamun_ruÎncă nu există evaluări
- 1 s2.0 S0092867410002369 MainDocument16 pagini1 s2.0 S0092867410002369 MainJames Matthew MadridÎncă nu există evaluări
- Cyberstalking and Cyberbullying in The Philippines LawDocument8 paginiCyberstalking and Cyberbullying in The Philippines LawJames Matthew MadridÎncă nu există evaluări
- ShibashiDocument3 paginiShibashiJames Matthew MadridÎncă nu există evaluări
- Hello WOrldDocument1 paginăHello WOrldJames Matthew MadridÎncă nu există evaluări
- Rubiks Cube 3x3 Solution-En PDFDocument13 paginiRubiks Cube 3x3 Solution-En PDFMidhun U K100% (1)
- Gantt ChartDocument3 paginiGantt ChartJames Matthew MadridÎncă nu există evaluări
- Gaudium Mass Songsheet Rev1Document2 paginiGaudium Mass Songsheet Rev1James Matthew MadridÎncă nu există evaluări
- TaizeDocument4 paginiTaizeJames Matthew Madrid100% (1)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Tet PGTRB Zoology Model Question Paper 2Document14 paginiTet PGTRB Zoology Model Question Paper 2Subbarayudu mamillaÎncă nu există evaluări
- Complete DentureDocument57 paginiComplete Denturedr_ahmad_zuhdi100% (3)
- Makabuhay IpDocument7 paginiMakabuhay IpButterflyCalmÎncă nu există evaluări
- Bharat India: Extremely Bad Status of Testing & VaccinationDocument308 paginiBharat India: Extremely Bad Status of Testing & VaccinationP Eng Suraj SinghÎncă nu există evaluări
- Jurnal GerdDocument6 paginiJurnal GerddilaÎncă nu există evaluări
- Mangosteen ManualDocument46 paginiMangosteen Manualherow999980% (5)
- 1st Internal Assessment Pharmacology 28.7.21 FinalDocument2 pagini1st Internal Assessment Pharmacology 28.7.21 FinalSubhom MajumderÎncă nu există evaluări
- MannitolDocument10 paginiMannitolFenzy Septiana AnggrainiÎncă nu există evaluări
- Basic First Aid TrainingDocument32 paginiBasic First Aid Trainingtukaram patilÎncă nu există evaluări
- Quality Issues of Milk & Milk Products (17-Uglc-603)Document22 paginiQuality Issues of Milk & Milk Products (17-Uglc-603)ayesha shafiqueÎncă nu există evaluări
- Bahasa Inggris UAN SMA 2010Document8 paginiBahasa Inggris UAN SMA 2010Abdullah RamliÎncă nu există evaluări
- An Overview of Mechanical Ventilation in The Intensive Care UnitDocument9 paginiAn Overview of Mechanical Ventilation in The Intensive Care UnitJaime Javier VegaÎncă nu există evaluări
- LamaDocument3 paginiLamarockyÎncă nu există evaluări
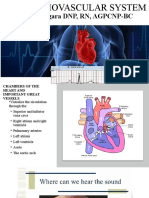
- The Cardiovascular System ReviewDocument18 paginiThe Cardiovascular System ReviewDanisha Reeves100% (1)
- Case Presentation OF: Iron Deficiency AnemiaDocument33 paginiCase Presentation OF: Iron Deficiency Anemiaitshurt_teardrops100% (1)
- Grape Growing in TennesseeDocument28 paginiGrape Growing in TennesseeDbaltÎncă nu există evaluări
- Indrabasti Marma2Document4 paginiIndrabasti Marma2Rajiv SharmaÎncă nu există evaluări
- Adrenal Function Urine TestDocument30 paginiAdrenal Function Urine TestDamarys ReyesÎncă nu există evaluări
- Aioh Position Paper DPM jdk2gdDocument26 paginiAioh Position Paper DPM jdk2gdRichardÎncă nu există evaluări
- Spiritual Wrestling PDFDocument542 paginiSpiritual Wrestling PDFJames CuasmayanÎncă nu există evaluări
- Clinical Findings and Management of PertussisDocument10 paginiClinical Findings and Management of PertussisAGUS DE COLSAÎncă nu există evaluări
- IOSH Module 1Document6 paginiIOSH Module 1Harbir Singh100% (1)
- Prune Belly SyndDocument11 paginiPrune Belly SyndnadiaryÎncă nu există evaluări
- Primary Health Care Notes Health What Is Illness and Causes of Illness?Document22 paginiPrimary Health Care Notes Health What Is Illness and Causes of Illness?Nano KaÎncă nu există evaluări
- Lesson 7 - Nervous, Endocrine, Reproductive, Urinary SystemDocument37 paginiLesson 7 - Nervous, Endocrine, Reproductive, Urinary SystemAlejandro GuerreroÎncă nu există evaluări
- Python Ieee Projects 2021 - 22 JPDocument3 paginiPython Ieee Projects 2021 - 22 JPWebsoft Tech-HydÎncă nu există evaluări
- DA Sistemik TerapiDocument11 paginiDA Sistemik TerapiariyatiÎncă nu există evaluări
- Urtica Urens William Boericke Cyrus Maxwell Bogar Adolf Zur Lippe John Henry ClarkeDocument9 paginiUrtica Urens William Boericke Cyrus Maxwell Bogar Adolf Zur Lippe John Henry ClarkeShah FaisalÎncă nu există evaluări
- Hypernatremia NEJM 2000Document8 paginiHypernatremia NEJM 2000BenjamÍn Alejandro Ruiz ManzanoÎncă nu există evaluări
- Family Health Survey ToolDocument7 paginiFamily Health Survey ToolReniella HidalgoÎncă nu există evaluări