S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- Dr. Hulda Clark - The Cure For All DiseasesDocument272 paginiDr. Hulda Clark - The Cure For All DiseasesRIS001100% (6)
- Nutrition and Infectious Diseases Shifting The Clinical Paradigm Humphries 1 Ed 2021Document524 paginiNutrition and Infectious Diseases Shifting The Clinical Paradigm Humphries 1 Ed 2021Anna Permyakova100% (1)
- Natural Science. Level 5. Unit 5. EcosystemsDocument4 paginiNatural Science. Level 5. Unit 5. EcosystemsAna Rosa Camacho Cornejo100% (1)
- Franklin 2005Document10 paginiFranklin 2005ahmrakÎncă nu există evaluări
- Effect of Live Yeast and Mannan-Oligosaccharides On Performance of Early-Lactation Holstein Dairy CowsDocument7 paginiEffect of Live Yeast and Mannan-Oligosaccharides On Performance of Early-Lactation Holstein Dairy CowsahmrakÎncă nu există evaluări
- FT42 Broz PDFDocument8 paginiFT42 Broz PDFahmrakÎncă nu există evaluări
- Effect of Supplementation of Enzyme Complex On Growth Performance of Crossbred PigsDocument5 paginiEffect of Supplementation of Enzyme Complex On Growth Performance of Crossbred PigsahmrakÎncă nu există evaluări
- A Practical Italian Approach To Controlling Aflatoxins in MilkDocument3 paginiA Practical Italian Approach To Controlling Aflatoxins in MilkahmrakÎncă nu există evaluări
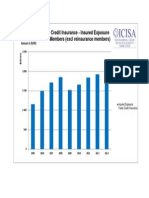
- Trade Credit Insurance - Insured Exposure ICISA Members (Excl Reinsurance Members)Document1 paginăTrade Credit Insurance - Insured Exposure ICISA Members (Excl Reinsurance Members)ahmrakÎncă nu există evaluări
- IAF - Radioökologie: 96132 SchlüsselfeldDocument1 paginăIAF - Radioökologie: 96132 SchlüsselfeldahmrakÎncă nu există evaluări
- DatabasevDocument2.288 paginiDatabasevahmrak100% (1)
- Effect of Betaine Source On Nutrient Utilisation of Broilers Challenged With CoccidiaDocument3 paginiEffect of Betaine Source On Nutrient Utilisation of Broilers Challenged With CoccidiaahmrakÎncă nu există evaluări
- Biological EvaluationDocument1 paginăBiological EvaluationahmrakÎncă nu există evaluări
- Aflatoxin in Dairy FeedsDocument4 paginiAflatoxin in Dairy FeedsahmrakÎncă nu există evaluări
- Pub 2897 Broiler PRDocument8 paginiPub 2897 Broiler PRahmrakÎncă nu există evaluări
- Supersect, ADWIA (Powder) : CompositionDocument1 paginăSupersect, ADWIA (Powder) : CompositionahmrakÎncă nu există evaluări
- A Dose Response Study With Lactylate in Broilers Challenged With A Subclinical Clostridium InfectionDocument1 paginăA Dose Response Study With Lactylate in Broilers Challenged With A Subclinical Clostridium InfectionahmrakÎncă nu există evaluări
- Egypt Non PasserineDocument9 paginiEgypt Non PasserineahmrakÎncă nu există evaluări
- Enzyme Linked Immunosorbent Assay ElisaDocument8 paginiEnzyme Linked Immunosorbent Assay ElisaAna Vitelariu - RaduÎncă nu există evaluări
- Ps 2007-00437Document9 paginiPs 2007-00437ahmrakÎncă nu există evaluări
- Folly Et Al, 2017 - Larvae Act As A Transient Transmission Hub For The Prevalent Bumblebee Parasite Crithidia BombiDocument5 paginiFolly Et Al, 2017 - Larvae Act As A Transient Transmission Hub For The Prevalent Bumblebee Parasite Crithidia BombiMaría Buendía AbadÎncă nu există evaluări
- Parasitology ReviewerDocument17 paginiParasitology Reviewerlouie100% (1)
- Parasitology Lab MaterialDocument43 paginiParasitology Lab MaterialFira'ol BogalaÎncă nu există evaluări
- Jss 2Document11 paginiJss 2Yusuf AbubakarÎncă nu există evaluări
- Relationships Among OrganismsDocument47 paginiRelationships Among OrganismsFirasat SultanÎncă nu există evaluări
- Parasites Behaviour and Welfare in FishDocument14 paginiParasites Behaviour and Welfare in FishRaúl GDÎncă nu există evaluări
- Answer Key Midterm 1 FRST395 For StudentsDocument5 paginiAnswer Key Midterm 1 FRST395 For StudentsGeorgia NunesÎncă nu există evaluări
- Parasitic InfectionsDocument27 paginiParasitic InfectionsYsabelle GutierrezÎncă nu există evaluări
- Species - Interaction - Worksheet AnswersDocument2 paginiSpecies - Interaction - Worksheet Answersnareman100% (1)
- The Effect of Learning and Search Images On Predator-Prey InteractionsDocument9 paginiThe Effect of Learning and Search Images On Predator-Prey Interactionsgaloli9055Încă nu există evaluări
- Terms and Concepts in Plant PathologyDocument48 paginiTerms and Concepts in Plant PathologytripsÎncă nu există evaluări
- HND Medical ScienceDocument588 paginiHND Medical Sciencembabit leslieÎncă nu există evaluări
- Bahasa Inggris Kel.8Document16 paginiBahasa Inggris Kel.8Egi Satya FralazendaÎncă nu există evaluări
- Efficacy of Herbal Ectoparasiticide Av/Epp/14 On Some Ectoparasites of Dogs and Cattle P.S. BanerjeeDocument3 paginiEfficacy of Herbal Ectoparasiticide Av/Epp/14 On Some Ectoparasites of Dogs and Cattle P.S. BanerjeeAmit Kumar PandeyÎncă nu există evaluări
- DLP Ecological RelationshipsDocument17 paginiDLP Ecological RelationshipsRuby Rose Molina100% (3)
- Biology Module Chap 8Document37 paginiBiology Module Chap 8Faida Hamid100% (4)
- Elementary Agriculture Teacher'S Notes Grade 5: Compiled By: TT Amadhila For Teachers in Oshikoto RegionDocument27 paginiElementary Agriculture Teacher'S Notes Grade 5: Compiled By: TT Amadhila For Teachers in Oshikoto RegionJB MangunduÎncă nu există evaluări
- The Full Moon Parasite Cleanse ProtocolDocument49 paginiThe Full Moon Parasite Cleanse ProtocolparthvarandaniÎncă nu există evaluări
- Learning Guide 05: Animal Health Care Service NTQF Level-IVDocument23 paginiLearning Guide 05: Animal Health Care Service NTQF Level-IVRafez Jone100% (1)
- Infestation: ScabiesDocument16 paginiInfestation: ScabiesBrix ValdrizÎncă nu există evaluări
- Microbiology and ParasitologyDocument42 paginiMicrobiology and ParasitologyJean Maraya100% (1)
- Review of LiteratureDocument11 paginiReview of LiteratureZahid NabiÎncă nu există evaluări
- TT PB 35 Lecture NotesDocument9 paginiTT PB 35 Lecture NotesAlex AndrangoÎncă nu există evaluări
- Interdependence Among Living Organisms and The EnviromentDocument20 paginiInterdependence Among Living Organisms and The EnviromentVisvan SubramaniamÎncă nu există evaluări
- Science-7 ECOLOGICAL RELATIONSHIPDocument5 paginiScience-7 ECOLOGICAL RELATIONSHIPBriansky Han100% (1)
- Lab CRPRDocument13 paginiLab CRPRdarryl calingacionÎncă nu există evaluări
- Companion Animal Parasitology: A Clinical Perspective: Peter J. IrwinDocument13 paginiCompanion Animal Parasitology: A Clinical Perspective: Peter J. IrwinAngela SanchezÎncă nu există evaluări