S-ar putea să vă placă și
- Eukaryotic Protein Synthesis Differs From Prokaryotic Protein Synthesis Primarily in Translation InitiationDocument4 paginiEukaryotic Protein Synthesis Differs From Prokaryotic Protein Synthesis Primarily in Translation InitiationJoyce DometitaÎncă nu există evaluări
- Protein Synthesis (Translation) - Microbiology - OpenStaxDocument7 paginiProtein Synthesis (Translation) - Microbiology - OpenStaxAleksandra Sanja MartinovicÎncă nu există evaluări
- Introduction To Translation: What You'll Learn To Do: Summarize The Process of TranslationDocument8 paginiIntroduction To Translation: What You'll Learn To Do: Summarize The Process of TranslationKempetsÎncă nu există evaluări
- BiochemDocument66 paginiBiochemReendelle JamiatreÎncă nu există evaluări
- Molecular Basis of Inheritance 2Document6 paginiMolecular Basis of Inheritance 2VARSHAN GAMINGÎncă nu există evaluări
- Protein Synthesis DefinitionDocument5 paginiProtein Synthesis Definitioncatherinebitkerrochester100% (2)
- Translation 190603145457Document30 paginiTranslation 190603145457soniaazizÎncă nu există evaluări
- Lecture 3: Translation and Mutations Learning GoalsDocument10 paginiLecture 3: Translation and Mutations Learning GoalsAngelica SmithÎncă nu există evaluări
- 4.central DogmaDocument4 pagini4.central DogmaAnanthan C.RÎncă nu există evaluări
- Chapter Fourteen: RNA Molecules and RNA Processing: Comprehension Questions Section 14.1Document11 paginiChapter Fourteen: RNA Molecules and RNA Processing: Comprehension Questions Section 14.1Qayyum LatipÎncă nu există evaluări
- SYBT Sem III Unit II TranslationDocument56 paginiSYBT Sem III Unit II TranslationMeir SabooÎncă nu există evaluări
- Essay Explaining The Process of Protein SynthesisDocument8 paginiEssay Explaining The Process of Protein SynthesisClaudia Acosta100% (1)
- Translation in ProkaryotesDocument25 paginiTranslation in ProkaryotesNaomi Lamtiur0% (1)
- BIO121 Tutorial - 07072021Document21 paginiBIO121 Tutorial - 07072021PARVATHY ANIL - IMS20211Încă nu există evaluări
- FALLSEM2023-24 BBIT307L TH VL2023240100207 2023-04-26 Reference-Material-IDocument29 paginiFALLSEM2023-24 BBIT307L TH VL2023240100207 2023-04-26 Reference-Material-IDrishti BisuiÎncă nu există evaluări
- CH 12Document69 paginiCH 12ᄋᄂᄋᄂÎncă nu există evaluări
- Kanzul Cell & Molecular BiologyDocument12 paginiKanzul Cell & Molecular BiologyMuhammad FarhanÎncă nu există evaluări
- Translation: From Messenger RNA To ProteinDocument71 paginiTranslation: From Messenger RNA To Proteinbombertest1Încă nu există evaluări
- Post Trasnscriptional Modification... ManuscriptDocument10 paginiPost Trasnscriptional Modification... ManuscriptAsif AhmedÎncă nu există evaluări
- MBII - L23 - Translation 2Document9 paginiMBII - L23 - Translation 2Miles NsgÎncă nu există evaluări
- Protein Synthesis in CellDocument7 paginiProtein Synthesis in Cellkvpyqegld100% (2)
- Protein Synthesis TranslationDocument5 paginiProtein Synthesis Translationrbafdvwhd100% (2)
- 2022-05-15 L8 - Protein SynthesisDocument64 pagini2022-05-15 L8 - Protein SynthesisTamara ElyasÎncă nu există evaluări
- Coursenotesdomain 4 InformationDocument29 paginiCoursenotesdomain 4 Informationapi-300668314Încă nu există evaluări
- TRANSLATIONDocument12 paginiTRANSLATIONNEET STUDIESÎncă nu există evaluări
- Protein Synthesis StepsDocument8 paginiProtein Synthesis Stepsgjga9bey100% (2)
- From Gene To Protein: Lecture OutlineDocument15 paginiFrom Gene To Protein: Lecture OutlineEiann Jasper LongcayanaÎncă nu există evaluări
- Utkarsh Sharma XII - A Biology Investigatory Project - TranslationDocument24 paginiUtkarsh Sharma XII - A Biology Investigatory Project - TranslationUtkarsh SharmaÎncă nu există evaluări
- Transcription &translation: Mol. Biology Lec-4 TranscriptionDocument12 paginiTranscription &translation: Mol. Biology Lec-4 TranscriptionAhmed Ali AssafÎncă nu există evaluări
- Protein Synthesis ProcessDocument4 paginiProtein Synthesis Processjamieakersfargo100% (1)
- De La Proteina La GenaDocument67 paginiDe La Proteina La GenaiuventasÎncă nu există evaluări
- InstructionsDocument5 paginiInstructionsLord NiquidÎncă nu există evaluări
- Molecular Final1Document9 paginiMolecular Final1h4hh9wqwvnÎncă nu există evaluări
- CHE 461 Module 5Document30 paginiCHE 461 Module 5Danladi VictoriaÎncă nu există evaluări
- Gene ExpressionDocument19 paginiGene Expressionkvicto100% (1)
- Resna N K MicrobiologyDocument54 paginiResna N K MicrobiologyResna N K ResiÎncă nu există evaluări
- BTN 332: Principles of Biotechnology 1 Name: Bobby Ikenna Asika Matric No: 18271070 Department: Microbiology Level: 300 AssignmentDocument1 paginăBTN 332: Principles of Biotechnology 1 Name: Bobby Ikenna Asika Matric No: 18271070 Department: Microbiology Level: 300 AssignmentGabriel SholaÎncă nu există evaluări
- AP Bio HW 17 091120Document4 paginiAP Bio HW 17 091120techchallenge2008Încă nu există evaluări
- Lec. 6Document10 paginiLec. 6Dr. Mohamed ShamsÎncă nu există evaluări
- Chapter IV Introduction To Bacterial Genetics-1Document67 paginiChapter IV Introduction To Bacterial Genetics-1temesgensemahegn55Încă nu există evaluări
- Genetic Memory PDFDocument5 paginiGenetic Memory PDFraba100Încă nu există evaluări
- Gene ExpressionDocument5 paginiGene ExpressionbenjaminlugahillaryÎncă nu există evaluări
- The Mechanism of TranslationDocument68 paginiThe Mechanism of TranslationRama KishoreÎncă nu există evaluări
- TranscriptionDocument25 paginiTranscriptionYamunaa ElencovanÎncă nu există evaluări
- BMB Test 3 Study GuidesDocument8 paginiBMB Test 3 Study GuidesNate LisbinÎncă nu există evaluări
- Apbio 17 LectureoutlineDocument16 paginiApbio 17 LectureoutlineGabe GallagherÎncă nu există evaluări
- From Gene To ProteinDocument16 paginiFrom Gene To ProteinPatricia NerÎncă nu există evaluări
- Protein BiosynthesisDocument21 paginiProtein BiosynthesisMyrrh Tagurigan TrainÎncă nu există evaluări
- 14 Solutions PDFDocument9 pagini14 Solutions PDFT CÎncă nu există evaluări
- CHAP 5 GeneticsDocument27 paginiCHAP 5 GeneticsAhmed AliÎncă nu există evaluări
- Protein Synthesis DiagramDocument6 paginiProtein Synthesis Diagramfygynejoheg2100% (1)
- Replication - Transcription - TranslationDocument75 paginiReplication - Transcription - TranslationJason FryÎncă nu există evaluări
- How Protien Is Being SynthesizedDocument5 paginiHow Protien Is Being SynthesizedMG Untalan BauzonÎncă nu există evaluări
- Gene Expression: Dr. Nor'Aishah HasanDocument57 paginiGene Expression: Dr. Nor'Aishah HasanLutfil HadiÎncă nu există evaluări
- Technological Institute of The Philippines 363 P. Casal ST., Quiapo, ManilaDocument6 paginiTechnological Institute of The Philippines 363 P. Casal ST., Quiapo, ManilaGellene GarciaÎncă nu există evaluări
- f215 Spec NotesDocument25 paginif215 Spec NoteswbaturleyÎncă nu există evaluări
- Chapter 6 GeneticsDocument12 paginiChapter 6 GeneticsAmer ToutonjiÎncă nu există evaluări
- Assessment Tool DuringDocument2 paginiAssessment Tool DuringEros Cuesta100% (1)
- Alexander Cummings: Mikrotoms (German For Description of A Microtome)Document1 paginăAlexander Cummings: Mikrotoms (German For Description of A Microtome)Eros CuestaÎncă nu există evaluări
- Role of Registered Nurses in Error Prevention, Discovery and CorrectionDocument4 paginiRole of Registered Nurses in Error Prevention, Discovery and CorrectionEros CuestaÎncă nu există evaluări
- Handover ERDocument12 paginiHandover EREros CuestaÎncă nu există evaluări
- Neonatal Jaundice NotesDocument5 paginiNeonatal Jaundice NotesEros CuestaÎncă nu există evaluări
- Africa 1Document3 paginiAfrica 1Eros CuestaÎncă nu există evaluări
- 06pain - 08Document51 pagini06pain - 08Vishwas U MadanÎncă nu există evaluări
- NurseDocument3 paginiNurseEros CuestaÎncă nu există evaluări
- BenedictsDocument12 paginiBenedictsEros CuestaÎncă nu există evaluări
- Drug StudyDocument5 paginiDrug StudyEros CuestaÎncă nu există evaluări
- NurseDocument3 paginiNurseEros CuestaÎncă nu există evaluări
- Go WorldDocument1 paginăGo WorldEros CuestaÎncă nu există evaluări
- Chemical Engineers Make Explosive ProgressDocument2 paginiChemical Engineers Make Explosive ProgressEros CuestaÎncă nu există evaluări
- MarketingDocument4 paginiMarketingEros CuestaÎncă nu există evaluări
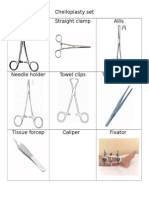
- Cheiloplasty SetDocument2 paginiCheiloplasty SetEros CuestaÎncă nu există evaluări
- Go WorldDocument1 paginăGo WorldEros CuestaÎncă nu există evaluări
- Goscript Reverse Transcriptase ProtocolDocument2 paginiGoscript Reverse Transcriptase Protocolinvestbiz optionstarÎncă nu există evaluări
- Iso 20395Document44 paginiIso 20395Gilberto Ragnar Nodarse100% (1)
- Salvia Divinorum Is A Highly EfficaciousDocument7 paginiSalvia Divinorum Is A Highly EfficaciousЗлобил ДружелюбовÎncă nu există evaluări
- Soliman (2018)Document12 paginiSoliman (2018)milaÎncă nu există evaluări
- Biotechnology AND It Application: By: Dr. Anand ManiDocument7 paginiBiotechnology AND It Application: By: Dr. Anand Mani....Încă nu există evaluări
- FERMENTAS Thermo-Scientific-Molecular-Biology-Workflow-Solutions-Brochure PDFDocument12 paginiFERMENTAS Thermo-Scientific-Molecular-Biology-Workflow-Solutions-Brochure PDFCarlos Arturo OsorioÎncă nu există evaluări
- BTH3752 Final Paper S2 2011Document4 paginiBTH3752 Final Paper S2 2011Grace ChoyÎncă nu există evaluări
- cDNA LibrariesDocument26 paginicDNA LibrariesRavi DesaiÎncă nu există evaluări
- Animal Trans Genesis and CloningDocument230 paginiAnimal Trans Genesis and CloningJanapareddi VenkataramanaÎncă nu există evaluări
- SOP For M PIMADocument9 paginiSOP For M PIMAk100% (1)
- Rediscovering Biology TextbookDocument203 paginiRediscovering Biology TextbookJuiced-IN itÎncă nu există evaluări
- Insight and Control of Infectious Disease in Global Scenario - P. Roy (Intech, 2012) WWDocument454 paginiInsight and Control of Infectious Disease in Global Scenario - P. Roy (Intech, 2012) WWgoutham sivasailamÎncă nu există evaluări
- Isolation of Intact Sub-Dermal Secretory Cavities From EucalyptusDocument11 paginiIsolation of Intact Sub-Dermal Secretory Cavities From EucalyptusManoakiÎncă nu există evaluări
- DG Midterm #1 Spring 2021 From ULA ReviewDocument7 paginiDG Midterm #1 Spring 2021 From ULA ReviewRITIKA SURISETTYÎncă nu există evaluări
- HIV Drug Resistance Training: Molecular Lab Set-Up and WorkflowDocument31 paginiHIV Drug Resistance Training: Molecular Lab Set-Up and WorkflowYantek Btklpp ManadoÎncă nu există evaluări
- Maxima H Minus First Strand cDNA Synthesis Kit: Thermo ScientificDocument2 paginiMaxima H Minus First Strand cDNA Synthesis Kit: Thermo ScientificEmily AguirreÎncă nu există evaluări
- Biotechnology in Healthcare - An Introduction To Biopharmaceuticals (PDFDrive)Document249 paginiBiotechnology in Healthcare - An Introduction To Biopharmaceuticals (PDFDrive)AddicoÎncă nu există evaluări
- Dr. M. A. Mazid Department of Pharmaceutical Chemistry, Faculty of Pharmacy, University of Dhaka, Dhaka-1000Document66 paginiDr. M. A. Mazid Department of Pharmaceutical Chemistry, Faculty of Pharmacy, University of Dhaka, Dhaka-100053-Deepankar SutradharÎncă nu există evaluări
- Advanced Diagnostic Aids in Oral PathologyDocument7 paginiAdvanced Diagnostic Aids in Oral PathologyIJAR JOURNALÎncă nu există evaluări
- Zeng (2017) - Reverse Genetic System of Bat SARS-like Coronaviruses and Function of ORFX - EnglishDocument92 paginiZeng (2017) - Reverse Genetic System of Bat SARS-like Coronaviruses and Function of ORFX - EnglishJose SilvaÎncă nu există evaluări
- 2016-Human Molecular Genetic and Functional Studies IdentifyDocument46 pagini2016-Human Molecular Genetic and Functional Studies IdentifyGabriel Heras ArribasÎncă nu există evaluări
- Immunohematology: Journal of Blood Group Serology and EducationDocument83 paginiImmunohematology: Journal of Blood Group Serology and Educationcodepalm650Încă nu există evaluări
- Growth HormoneDocument7 paginiGrowth Hormonefs semjohnÎncă nu există evaluări
- Gel Electrophoresis Lab ReportDocument5 paginiGel Electrophoresis Lab ReportAngela Leong Feng PingÎncă nu există evaluări
- Cancers 14 01516Document20 paginiCancers 14 01516alexcento74Încă nu există evaluări
- Genetic EngineeringDocument27 paginiGenetic EngineeringSeema QureshiÎncă nu există evaluări
- Cloning StrategiesDocument35 paginiCloning Strategiesskmishra8467% (3)
- Cdna and Bac LibraryDocument14 paginiCdna and Bac LibraryNoman AnwarÎncă nu există evaluări
- Revised TERM PAPERDocument19 paginiRevised TERM PAPERVikal RajputÎncă nu există evaluări
- Disease MarkersDocument91 paginiDisease MarkersAlbertina MontenegroÎncă nu există evaluări
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDe la EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisEvaluare: 3.5 din 5 stele3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDe la EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceEvaluare: 4.5 din 5 stele4.5/5 (516)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDe la EverandTales from Both Sides of the Brain: A Life in NeuroscienceEvaluare: 3 din 5 stele3/5 (18)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)De la EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Evaluare: 4 din 5 stele4/5 (378)
- Gut: the new and revised Sunday Times bestsellerDe la EverandGut: the new and revised Sunday Times bestsellerEvaluare: 4 din 5 stele4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDe la EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityEvaluare: 4 din 5 stele4/5 (3)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDe la EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsEvaluare: 4.5 din 5 stele4.5/5 (6)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDe la EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouEvaluare: 4.5 din 5 stele4.5/5 (62)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDe la EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeÎncă nu există evaluări
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDe la EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperEvaluare: 4.5 din 5 stele4.5/5 (15)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDe la EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesEvaluare: 4.5 din 5 stele4.5/5 (397)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDe la EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionEvaluare: 4 din 5 stele4/5 (811)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldDe la EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldEvaluare: 4 din 5 stele4/5 (595)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDe la Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessEvaluare: 4 din 5 stele4/5 (33)
- Who's in Charge?: Free Will and the Science of the BrainDe la EverandWho's in Charge?: Free Will and the Science of the BrainEvaluare: 4 din 5 stele4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDe la EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorÎncă nu există evaluări
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDe la EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindEvaluare: 4.5 din 5 stele4.5/5 (93)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDe la EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedEvaluare: 4 din 5 stele4/5 (11)
- Good Without God: What a Billion Nonreligious People Do BelieveDe la EverandGood Without God: What a Billion Nonreligious People Do BelieveEvaluare: 4 din 5 stele4/5 (66)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDe la EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemEvaluare: 4.5 din 5 stele4.5/5 (115)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDe la EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldEvaluare: 4.5 din 5 stele4.5/5 (18)
- Seven and a Half Lessons About the BrainDe la EverandSeven and a Half Lessons About the BrainEvaluare: 4 din 5 stele4/5 (109)
- Human: The Science Behind What Makes Your Brain UniqueDe la EverandHuman: The Science Behind What Makes Your Brain UniqueEvaluare: 3.5 din 5 stele3.5/5 (38)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDe la EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildEvaluare: 4.5 din 5 stele4.5/5 (44)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessDe la EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessEvaluare: 4 din 5 stele4/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDe la EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondEvaluare: 4 din 5 stele4/5 (4)