S-ar putea să vă placă și
- Funcion de Los Aminoacidos Como BioestimulantesDocument3 paginiFuncion de Los Aminoacidos Como BioestimulantesRudy Puma VilcaÎncă nu există evaluări
- Fitotoxicidad de Cobre en Lechuga Unh PDFDocument8 paginiFitotoxicidad de Cobre en Lechuga Unh PDFCesarÎncă nu există evaluări
- Artículo Del Frijol. Jesús Montúfar.Document16 paginiArtículo Del Frijol. Jesús Montúfar.MontúfarÎncă nu există evaluări
- Clase UNIDAD 7 Herenc CitoplDocument45 paginiClase UNIDAD 7 Herenc CitoplEmilio AbdalaÎncă nu există evaluări
- Cálculo de La Solución Nutritiva en Fertirriego. General o EstándarDocument6 paginiCálculo de La Solución Nutritiva en Fertirriego. General o EstándarRómulo Del ValleÎncă nu există evaluări
- Ciampitti y Garcia 2007b PDFDocument4 paginiCiampitti y Garcia 2007b PDFDerly LeonÎncă nu există evaluări
- Aminoacidos en La AgriculturaDocument11 paginiAminoacidos en La AgriculturaRoanmy Schol ChalibÎncă nu există evaluări
- Prácticas de Laboratorio de Proteccion VegetalDocument56 paginiPrácticas de Laboratorio de Proteccion VegetalAndy Tomala100% (1)
- Solución Nutritiva para El TomateDocument3 paginiSolución Nutritiva para El TomateRotciv Asos ZelaznogÎncă nu există evaluări
- Curvas de absorción de nutrientes en papaya PococíDocument12 paginiCurvas de absorción de nutrientes en papaya PococíAntonio AlvaradoÎncă nu există evaluări
- Bacillus SporothermoduransDocument4 paginiBacillus SporothermoduransLuisMiguelBolañosÎncă nu există evaluări
- Ecofisiología de Coliflor PDFDocument65 paginiEcofisiología de Coliflor PDFPao GabrielÎncă nu există evaluări
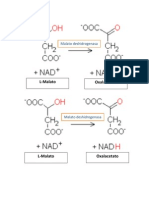
- La Enzima Malato DeshidrogenasaDocument2 paginiLa Enzima Malato DeshidrogenasaKaren Torres JuárezÎncă nu există evaluări
- TriacontanolDocument32 paginiTriacontanolcristiam leguia yaurisÎncă nu există evaluări
- Beta Glucanos Y Beta GlucanasasDocument19 paginiBeta Glucanos Y Beta GlucanasasbirrashoppÎncă nu există evaluări
- CitoquiminaDocument6 paginiCitoquiminamarcelino canchari caceresÎncă nu există evaluări
- Ciclo de CalvinDocument58 paginiCiclo de CalvinAriana Puicon100% (1)
- Deficiencias y Excesos Nutricionales en TomateDocument25 paginiDeficiencias y Excesos Nutricionales en TomatemiguelÎncă nu există evaluări
- Enfermedades en Tomate Bajo InvernaderoDocument54 paginiEnfermedades en Tomate Bajo InvernaderoDanielÎncă nu există evaluări
- Recomendaciones para El TomateDocument40 paginiRecomendaciones para El TomateJose Luis Colin MarinezÎncă nu există evaluări
- Fertilidad de SuelosDocument37 paginiFertilidad de SuelosRolling Richard Loayza Fernandez100% (3)
- 22 Viroides PDFDocument11 pagini22 Viroides PDFThewilson H. RamosÎncă nu există evaluări
- Las Micorrizas y La Absorción Del FósforoDocument2 paginiLas Micorrizas y La Absorción Del FósforoIlse MireyaÎncă nu există evaluări
- Comentario de ArmadansDocument2 paginiComentario de ArmadansgermanÎncă nu există evaluări
- Actividad enzimática peroxidasaDocument27 paginiActividad enzimática peroxidasaJavier ClarosÎncă nu există evaluări
- Protocolo Bioestimulantes CoagronorteDocument9 paginiProtocolo Bioestimulantes CoagronorteMultiservicios E&EÎncă nu există evaluări
- Giberelinas 2016Document72 paginiGiberelinas 2016MariaÎncă nu există evaluări
- CultivoDocument7 paginiCultivoCarolina Panchana TÎncă nu există evaluări
- Factores Que Afectan La FotosintesisDocument16 paginiFactores Que Afectan La FotosintesisLitzy ValenzuelaÎncă nu există evaluări
- Memorias FruticulturaDocument100 paginiMemorias FruticulturaMario Jose Yovera ReyesÎncă nu există evaluări
- Fi Totec NiaDocument224 paginiFi Totec NiamartaÎncă nu există evaluări
- Funciones carbohidratos organismos vivos clasificación enlacesDocument29 paginiFunciones carbohidratos organismos vivos clasificación enlacesyemeneses126Încă nu există evaluări
- Micropropagación en RosasDocument9 paginiMicropropagación en RosasGuido Sanchez RojasÎncă nu există evaluări
- AguacateDocument44 paginiAguacatejamiro52Încă nu există evaluări
- Investigacion Sobre El MangoDocument30 paginiInvestigacion Sobre El MangoDeilis HerreraÎncă nu există evaluări
- Estrategia Fertil I Zac I On Vid de MesaDocument40 paginiEstrategia Fertil I Zac I On Vid de Mesafvasquez25Încă nu există evaluări
- Desinfestación de sustratos y soluciones nutritivas contaminadas con Fusarium oxysporum f. sp. lycopersici en cultivo de tomate sin sueloDocument71 paginiDesinfestación de sustratos y soluciones nutritivas contaminadas con Fusarium oxysporum f. sp. lycopersici en cultivo de tomate sin sueloManu AmaroÎncă nu există evaluări
- Uso de Aminoácidos en La AgriculturaDocument4 paginiUso de Aminoácidos en La AgriculturaHéctor AlfonsoÎncă nu există evaluări
- Citrus Tristeza Virus - 2011 PDFDocument23 paginiCitrus Tristeza Virus - 2011 PDFEdgar Couttolenc BrenisÎncă nu există evaluări
- Via Del Acido ShiquimicoDocument29 paginiVia Del Acido ShiquimicoAnonymous cOnoOsmSSPÎncă nu există evaluări
- 2 - Origen MaizDocument35 pagini2 - Origen MaizAlexander Oviedo BenítezÎncă nu există evaluări
- 2 Examen de Fisiología Vegetal AvanzadaDocument3 pagini2 Examen de Fisiología Vegetal AvanzadaNormaÎncă nu există evaluări
- B. Bassiana en ArrozDocument78 paginiB. Bassiana en ArrozmorenoandyÎncă nu există evaluări
- Agrometeorología: Conceptos fundamentales del clima y su impacto en la agriculturaDocument39 paginiAgrometeorología: Conceptos fundamentales del clima y su impacto en la agriculturaBerlin Saimon100% (1)
- Taxonomía de Los HongosDocument11 paginiTaxonomía de Los HongosVioleta Lazaro JimenezÎncă nu există evaluări
- Xilotrom Es IpDocument20 paginiXilotrom Es IpCarlos Augusto Alvarez LondoñoÎncă nu există evaluări
- Curva de crecimiento simple sigmoidal de la fresaDocument7 paginiCurva de crecimiento simple sigmoidal de la fresaPepe MejiaÎncă nu există evaluări
- Mite KillDocument3 paginiMite KillSebastian QuiñonesÎncă nu există evaluări
- El Receso en FrutalesDocument12 paginiEl Receso en FrutalesGabriela VilchesÎncă nu există evaluări
- Anthoxanthum OdoratumDocument1 paginăAnthoxanthum OdoratumVictor Alexis Avendaño RodriguezÎncă nu există evaluări
- Tipos de CitocininasDocument5 paginiTipos de CitocininasAlfonso Cadenas LemusÎncă nu există evaluări
- Funciones del boro en la plantaDocument4 paginiFunciones del boro en la plantagonzalo_hbÎncă nu există evaluări
- Fisiologia VegetalDocument11 paginiFisiologia Vegetalcarlostk16Încă nu există evaluări
- El Blog de Jair Beltrán - Informe Practica de Imbibición (Fisiología Vegetal)Document7 paginiEl Blog de Jair Beltrán - Informe Practica de Imbibición (Fisiología Vegetal)frankoÎncă nu există evaluări
- Practica de Los FrijolesDocument10 paginiPractica de Los FrijolesLiliana Alavez SosaÎncă nu există evaluări
- Manejo fitosanitario del cultivo de hortalizas medidas para la temporada invernalDe la EverandManejo fitosanitario del cultivo de hortalizas medidas para la temporada invernalÎncă nu există evaluări
- Algunas especies de moscas de la fruta, exóticas para ColombiaDe la EverandAlgunas especies de moscas de la fruta, exóticas para ColombiaÎncă nu există evaluări
- Trabajo de BiologíaDocument12 paginiTrabajo de BiologíaMaria Pia SánchezÎncă nu există evaluări
- Características Clínicas de La Enfermedad Por Coronavirus 2019 en ChinaDocument13 paginiCaracterísticas Clínicas de La Enfermedad Por Coronavirus 2019 en ChinaCristian Moreno CamañoÎncă nu există evaluări
- Dinf PC03 08Document1 paginăDinf PC03 08mayra alejandraÎncă nu există evaluări
- Características Clínicas de La Enfermedad Por Coronavirus 2019 en ChinaDocument13 paginiCaracterísticas Clínicas de La Enfermedad Por Coronavirus 2019 en ChinaCristian Moreno CamañoÎncă nu există evaluări
- Genetic Admixture Estimates by Alu Elements in Afro-Colombian and Mestizo Populations From Antioquia, Colombia 2010Document13 paginiGenetic Admixture Estimates by Alu Elements in Afro-Colombian and Mestizo Populations From Antioquia, Colombia 2010Cristian Moreno CamañoÎncă nu există evaluări
- Frente A Covid-19 en Italia Ética, Logística y Terapéutica en La Primera Línea de La EpidemiaDocument6 paginiFrente A Covid-19 en Italia Ética, Logística y Terapéutica en La Primera Línea de La EpidemiaCristian Moreno CamañoÎncă nu există evaluări
- La Autentificación Biométrica Tiene Como FinalidadDocument4 paginiLa Autentificación Biométrica Tiene Como FinalidadCristian Moreno CamañoÎncă nu există evaluări
- 03 LípidosDocument7 pagini03 LípidosbeabergamoÎncă nu există evaluări
- Escapar de La Caja de PandoraDocument5 paginiEscapar de La Caja de PandoraCristian Moreno CamañoÎncă nu există evaluări
- El Triaje Más Duro - Asignación de Ventiladores en Una PandemiaDocument4 paginiEl Triaje Más Duro - Asignación de Ventiladores en Una PandemiaCristian Moreno CamañoÎncă nu există evaluări
- Actividades Rompehielo PDFDocument14 paginiActividades Rompehielo PDFvladiÎncă nu există evaluări
- Im Spanish K-Mid100 09-03-2017Document12 paginiIm Spanish K-Mid100 09-03-2017Cristian Moreno CamañoÎncă nu există evaluări
- Joven conforme al corazón de DiosDocument4 paginiJoven conforme al corazón de DiosCristian Moreno Camaño67% (3)
- Conservación Ex Situ Caso Del Jaguar yDocument1 paginăConservación Ex Situ Caso Del Jaguar yCristian Moreno CamañoÎncă nu există evaluări
- Dones EspiritualesDocument75 paginiDones EspiritualesCristian Moreno CamañoÎncă nu există evaluări
- Ciudades como ecosistemas: flujos de energía y materiaDocument2 paginiCiudades como ecosistemas: flujos de energía y materiaCristian Moreno CamañoÎncă nu există evaluări
- AcneDocument289 paginiAcnechicho6404100% (2)
- Antopodos-equinodermos-Anelidos de Bahia CispataDocument11 paginiAntopodos-equinodermos-Anelidos de Bahia CispataCristian Moreno CamañoÎncă nu există evaluări
- Carbon oDocument3 paginiCarbon oCristian Moreno CamañoÎncă nu există evaluări
- AlvaDocument21 paginiAlvaCristian Moreno CamañoÎncă nu există evaluări
- Nombres CientificosDocument2 paginiNombres CientificosCristian Moreno CamañoÎncă nu există evaluări
- Taller Molecular 1Document2 paginiTaller Molecular 1Cristian Moreno Camaño0% (2)
- Diapositivas Informe MosquitoDocument10 paginiDiapositivas Informe MosquitoCristian Moreno CamañoÎncă nu există evaluări
- 2 Morfo Tallo Raiz 1Document17 pagini2 Morfo Tallo Raiz 1Cristian Moreno Camaño100% (1)
- Mutaciones Cromosomicas Estructurales Miguel-PitaDocument60 paginiMutaciones Cromosomicas Estructurales Miguel-PitaCristian Moreno CamañoÎncă nu există evaluări
- Morfologia TalloDocument16 paginiMorfologia TalloAngeles RvÎncă nu există evaluări
- ALGASDocument2 paginiALGASCristian Moreno CamañoÎncă nu există evaluări
- Adn MosquitoDocument4 paginiAdn MosquitoCristian Moreno CamañoÎncă nu există evaluări
- Familia JuncaceaeDocument10 paginiFamilia JuncaceaeCristian Moreno CamañoÎncă nu există evaluări
- Bahia de CsipatáDocument8 paginiBahia de CsipatáCristian Moreno CamañoÎncă nu există evaluări
- PROPAGANDADocument3 paginiPROPAGANDAAndres HerreraÎncă nu există evaluări
- 180 Face WashDocument3 pagini180 Face WashCarolina ParzÎncă nu există evaluări
- Trabajo de Propoleo en Presentacion en Power PointDocument60 paginiTrabajo de Propoleo en Presentacion en Power PointRigo Lozada100% (1)
- Te Hindu PDFDocument22 paginiTe Hindu PDFMaria ColqueÎncă nu există evaluări
- InhibmodifDocument46 paginiInhibmodifLuis SanchezÎncă nu există evaluări
- Vida Sin Gastritis PDF GratisDocument7 paginiVida Sin Gastritis PDF GratisRene LopezÎncă nu există evaluări
- Enciendetupineal PDFDocument30 paginiEnciendetupineal PDFLucas Thomas100% (3)
- Oxidación frutasDocument5 paginiOxidación frutasseeb videosÎncă nu există evaluări
- Proyecto Final PRQ 207Document101 paginiProyecto Final PRQ 207Da NiÎncă nu există evaluări
- Presentacion Bear BerryDocument13 paginiPresentacion Bear BerryPaly de HuifeÎncă nu există evaluări
- Articulos 2012 (Fitness Revolucionario) PDFDocument387 paginiArticulos 2012 (Fitness Revolucionario) PDFMaxi FogliaÎncă nu există evaluări
- Go Well PresentacionDocument13 paginiGo Well PresentacionSandra Valeria Roncal GarcíaÎncă nu există evaluări
- 19 Beneficios saludables de la cebollaDocument8 pagini19 Beneficios saludables de la cebollarosa carraraÎncă nu există evaluări
- Catalogo ProfesionalDocument14 paginiCatalogo ProfesionalAlex LlanosÎncă nu există evaluări
- Sachapapa MoradaDocument3 paginiSachapapa MoradaYodiOlazaÎncă nu există evaluări
- Presentacion de Bebidas DXNDocument28 paginiPresentacion de Bebidas DXNCyber BluestarÎncă nu există evaluări
- Tea AlimentaciónDocument26 paginiTea AlimentaciónXIOMARA DEL CARMEN BARRANZUELA NEYRAÎncă nu există evaluări
- Cromo y Suplementos de Yodo y La Baja de PesoDocument9 paginiCromo y Suplementos de Yodo y La Baja de PesoAnonymous Yw70EVHkeÎncă nu există evaluări
- Contenido de Olla AlimentariaDocument20 paginiContenido de Olla AlimentariaCésar Hernández100% (1)
- Comidas InfelicesDocument21 paginiComidas Infelicesarmado421Încă nu există evaluări
- Para Cuaderno de CampoDocument7 paginiPara Cuaderno de CamposamiuÎncă nu există evaluări
- Catalogo Fuxion Col PDFDocument56 paginiCatalogo Fuxion Col PDFDavid Mauricio Valencia ClarosÎncă nu există evaluări
- PLÁTANODocument3 paginiPLÁTANOMATA_CRITTERSÎncă nu există evaluări
- Sanky: antioxidante andino de alto valorDocument2 paginiSanky: antioxidante andino de alto valorandreaÎncă nu există evaluări
- Propiedades de La SabilaDocument17 paginiPropiedades de La SabilaRoberto GonzalezÎncă nu există evaluări
- Vida Cápsulas ingredientes para perder peso de forma saludableDocument4 paginiVida Cápsulas ingredientes para perder peso de forma saludableMarcelo SalasÎncă nu există evaluări
- Determinación de Vitamina CDocument3 paginiDeterminación de Vitamina CElvia Roxana Guzmán GarcíaÎncă nu există evaluări
- Lamina 1 PDFDocument2 paginiLamina 1 PDFKatyyCaucamanÎncă nu există evaluări
- Boddy ButtersDocument39 paginiBoddy ButtersCeip Isabel Fajardo100% (2)