S-ar putea să vă placă și
- Patch Testing and Prick Testing: A Practical Guide Official Publication of the ICDRGDe la EverandPatch Testing and Prick Testing: A Practical Guide Official Publication of the ICDRGÎncă nu există evaluări
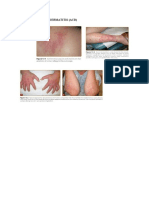
- Irritant Contact Dermatitis: Causes, Symptoms, TreatmentDocument13 paginiIrritant Contact Dermatitis: Causes, Symptoms, TreatmentYoga Alfian NoorÎncă nu există evaluări
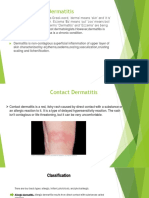
- Contact DermatitisDocument4 paginiContact DermatitisErlangga D SaputroÎncă nu există evaluări
- Calixtro, LJ Contact Dermatitis NarrativeDocument2 paginiCalixtro, LJ Contact Dermatitis NarrativeKim SunooÎncă nu există evaluări
- Bercak MerahDocument16 paginiBercak MerahFary SatriadiÎncă nu există evaluări
- DermaDocument9 paginiDermaZain Ul AbidinÎncă nu există evaluări
- Irritant Contact DermatitisDocument5 paginiIrritant Contact DermatitisJohn P M SinagaÎncă nu există evaluări
- Dermatitis: K. Puviyarasi Dept of MSN MNCDocument26 paginiDermatitis: K. Puviyarasi Dept of MSN MNCPuviyarasi100% (2)
- Contact DermatitisDocument2 paginiContact DermatitisybhuiaÎncă nu există evaluări
- 334-Article Text-1213-1-10-20160514Document7 pagini334-Article Text-1213-1-10-20160514Balakrishna GopinathÎncă nu există evaluări
- Toxic Responses of The SkinDocument25 paginiToxic Responses of The SkinfianceeleeÎncă nu există evaluări
- Allergic Skin Conditions - Causes, Clinical Features and TreatmentDocument4 paginiAllergic Skin Conditions - Causes, Clinical Features and TreatmentAnshuman PadheeÎncă nu există evaluări
- Contact Dermatitis in ChildrenDocument6 paginiContact Dermatitis in ChildrenYogi SanjayaÎncă nu există evaluări
- Pathophysiological Differences Between Irritant and Allergic Contact DermatitisDocument8 paginiPathophysiological Differences Between Irritant and Allergic Contact DermatitisFenni OktoberryÎncă nu există evaluări
- Contact Dermatitis: Key PointsDocument8 paginiContact Dermatitis: Key Pointssaimon reyÎncă nu există evaluări
- Eczema and DermatitisDocument24 paginiEczema and Dermatitisjackofmanytrades100% (1)
- Case Report Dermatitis ContactDocument10 paginiCase Report Dermatitis ContactMuhammad Aktora Tarigan100% (1)
- Contact Dermatitis Contact Dermatitis Is A Term For A Skin Reaction (Dermatitis) Resulting From Exposure To AllergensDocument4 paginiContact Dermatitis Contact Dermatitis Is A Term For A Skin Reaction (Dermatitis) Resulting From Exposure To AllergensAlbert Simon GalizaÎncă nu există evaluări
- Synopsis of Causation: Dermatitis (Eczema) Including Occupational DermatitisDocument19 paginiSynopsis of Causation: Dermatitis (Eczema) Including Occupational DermatitisCcÎncă nu există evaluări
- Acute Inflammatory DermatosesDocument29 paginiAcute Inflammatory DermatosesHaniya KhanÎncă nu există evaluări
- Dermatitis Kontak Alergi (DKA)Document25 paginiDermatitis Kontak Alergi (DKA)tryhabibullahÎncă nu există evaluări
- Referensi 7Document9 paginiReferensi 7iffaÎncă nu există evaluări
- Contact dermatitis rash causesDocument31 paginiContact dermatitis rash causesBrendaÎncă nu există evaluări
- Toxic Responses of the Skin: Skin Functions and DiseasesDocument6 paginiToxic Responses of the Skin: Skin Functions and DiseasesM. JoyceÎncă nu există evaluări
- ms24 PDFDocument8 paginims24 PDFDimas F. HidayatÎncă nu există evaluări
- Allergic Contact DermatitisDocument11 paginiAllergic Contact DermatitisSf AkhadiyatiÎncă nu există evaluări
- 2 EczemaDocument5 pagini2 Eczemaحسين طاهر حاتم طاهرÎncă nu există evaluări
- Veterinary Dermatitis ReportDocument11 paginiVeterinary Dermatitis ReportJoey LwÎncă nu există evaluări
- Allergic Contact Dermatitis - TranslateDocument9 paginiAllergic Contact Dermatitis - Translatewahyuni taslimÎncă nu există evaluări
- Generalized Eczematous Rash in A Patient With Shoe Contact Dermatitis A Case ReportDocument3 paginiGeneralized Eczematous Rash in A Patient With Shoe Contact Dermatitis A Case Reportcgs08Încă nu există evaluări
- DddsDocument50 paginiDddsOsama AlhaseÎncă nu există evaluări
- Dermatitis and Eczema: Ailing Zou (邹爱玲) 2021.10.18Document31 paginiDermatitis and Eczema: Ailing Zou (邹爱玲) 2021.10.18AmeliaÎncă nu există evaluări
- Journal of Allergy & TherapyDocument8 paginiJournal of Allergy & TherapyseluarbolaÎncă nu există evaluări
- Contact Dermatitis. The Major Occupational Allergic Skin DiseasesDocument66 paginiContact Dermatitis. The Major Occupational Allergic Skin DiseasesJulius HabibiÎncă nu există evaluări
- Allergic Contact Dermatitis Mechanisms ReviewedDocument11 paginiAllergic Contact Dermatitis Mechanisms ReviewedngekÎncă nu există evaluări
- Skin Disorders Guide: Urticaria, Eczema, Contact DermatitisDocument15 paginiSkin Disorders Guide: Urticaria, Eczema, Contact DermatitisManar AlzobiÎncă nu există evaluări
- Irritant Contact Dermatitis from Cosmetic Soap (39chDocument10 paginiIrritant Contact Dermatitis from Cosmetic Soap (39chstillmysunsetÎncă nu există evaluări
- Contact DDocument12 paginiContact DSeenha DewanÎncă nu există evaluări
- Adult Nursing 2020/ Integumentary System Assist Lect /shelan Qahraman SkinDocument8 paginiAdult Nursing 2020/ Integumentary System Assist Lect /shelan Qahraman Skinمحمد سعد طهÎncă nu există evaluări
- WEEK 8 - Skincare Products Part 2Document31 paginiWEEK 8 - Skincare Products Part 2johnbiel.barsana26Încă nu există evaluări
- Identifying The Cause of Contact DermatitisDocument6 paginiIdentifying The Cause of Contact DermatitiswadejackÎncă nu există evaluări
- Case ReportDocument13 paginiCase ReportmarselamgeÎncă nu există evaluări
- Allergy and HypersensitivityDocument49 paginiAllergy and HypersensitivityMoshe Cohen'sÎncă nu există evaluări
- RetinoidDocument22 paginiRetinoidPresstisa AxeliaÎncă nu există evaluări
- Dermatitis & Urticaria: Department of Dermato Venereology Faculty of Medicine Gadjah Mada UniversityDocument34 paginiDermatitis & Urticaria: Department of Dermato Venereology Faculty of Medicine Gadjah Mada UniversityadystiÎncă nu există evaluări
- Cutaneous Manifestation of Toxic ExposuresDocument41 paginiCutaneous Manifestation of Toxic Exposureserrol.williamson19Încă nu există evaluări
- Mechanisms of Inflammation I, II, and III LOsDocument9 paginiMechanisms of Inflammation I, II, and III LOsAndrew SagalovÎncă nu există evaluări
- Dermatitis KuliahDocument17 paginiDermatitis KuliahFirda LudfiyahÎncă nu există evaluări
- HypersensitivityDocument49 paginiHypersensitivityAkanksha MahajanÎncă nu există evaluări
- Penyakit Dermatitis: Dr. Kristo A. Nababan, SPKKDocument47 paginiPenyakit Dermatitis: Dr. Kristo A. Nababan, SPKKDaniel Suarez ParapatÎncă nu există evaluări
- Xerosis CutisDocument6 paginiXerosis CutisHadi FirmansyahÎncă nu există evaluări
- Contact Dermatitis BJD Guidelines May 2009Document9 paginiContact Dermatitis BJD Guidelines May 2009Cynthia OktariszaÎncă nu există evaluări
- Hand DermatitisDocument6 paginiHand Dermatitisminerva_stanciuÎncă nu există evaluări
- Dermatitis: Department of Dermato Venereology Faculty of Medicine Gadjah Mada UniversityDocument30 paginiDermatitis: Department of Dermato Venereology Faculty of Medicine Gadjah Mada UniversityMuhammad mukramÎncă nu există evaluări
- Dermatitis Kuliah OpDocument17 paginiDermatitis Kuliah OpTrii SakttiÎncă nu există evaluări
- Presentation PhytoDocument57 paginiPresentation Phytoyasmeenelraouf16Încă nu există evaluări
- Ectoin AntiinflamasiDocument19 paginiEctoin AntiinflamasiSiti fatimahÎncă nu există evaluări
- Journal Reading Kulit - Nixal Kurniawan (En)Document18 paginiJournal Reading Kulit - Nixal Kurniawan (En)Nixal KurniawanÎncă nu există evaluări
- Ijirt161480 PaperDocument11 paginiIjirt161480 PaperCIVILERGAURAVVERMAÎncă nu există evaluări
- Ns. M. Nurman, M. KepDocument30 paginiNs. M. Nurman, M. KepSabaniaÎncă nu există evaluări
- Satria Adi PutraDocument83 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- DXA in Adults: The Challenge of DiagnosisDocument4 paginiDXA in Adults: The Challenge of DiagnosisSatria Adi PutraÎncă nu există evaluări
- Satria AdiDocument5 paginiSatria AdiSatria Adi PutraÎncă nu există evaluări
- Basic Principles and Technical Considerations: Radiographic AbsorptiometryDocument10 paginiBasic Principles and Technical Considerations: Radiographic AbsorptiometrySatria Adi PutraÎncă nu există evaluări
- Satria AdiDocument10 paginiSatria AdiSatria Adi PutraÎncă nu există evaluări
- Satria AdiDocument10 paginiSatria AdiSatria Adi PutraÎncă nu există evaluări
- Satria AdiDocument13 paginiSatria AdiSatria Adi PutraÎncă nu există evaluări
- Bed Positioning For Stroke Patients: A. Lying On Your Back or SittingDocument1 paginăBed Positioning For Stroke Patients: A. Lying On Your Back or SittingSatria Adi PutraÎncă nu există evaluări
- Alternative Ways To Retrieved Articles From The Internet (Useful Information)Document8 paginiAlternative Ways To Retrieved Articles From The Internet (Useful Information)Agus SudarmaÎncă nu există evaluări
- Satria Adi PutraDocument3 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- Satria Adi PutraDocument6 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- Satria Adi PutraDocument83 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- Satria Adi PutraDocument11 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- Satria Adi PutraDocument5 paginiSatria Adi PutraSatria Adi PutraÎncă nu există evaluări
- Paper Template For 2nd ASEAN Academic Society International ConferenceDocument3 paginiPaper Template For 2nd ASEAN Academic Society International ConferenceSatria Adi PutraÎncă nu există evaluări
- DratDocument1 paginăDratSatria Adi PutraÎncă nu există evaluări
- Article DHEA and AntiagingDocument2 paginiArticle DHEA and AntiagingSatria Adi PutraÎncă nu există evaluări
- Answers 19521Document845 paginiAnswers 19521docerick87Încă nu există evaluări
- LearnEnglish Reading B2 A Plastic Ocean A Film ReviewDocument5 paginiLearnEnglish Reading B2 A Plastic Ocean A Film ReviewSerly FimasariÎncă nu există evaluări
- Managing Mental Health and Stress Online CourseDocument2 paginiManaging Mental Health and Stress Online CourseDakota624Încă nu există evaluări
- OSHS LegislationsDocument59 paginiOSHS LegislationsAlvin DeliroÎncă nu există evaluări
- People Pleasing Patterns Are Learned When Needs Are Not Met PDFDocument10 paginiPeople Pleasing Patterns Are Learned When Needs Are Not Met PDFPamela RodriguezÎncă nu există evaluări
- Hospital Administrative Assistant Resume-A4Document2 paginiHospital Administrative Assistant Resume-A4Philip V AlexÎncă nu există evaluări
- Elanco Parvovirus DXTX GDocument2 paginiElanco Parvovirus DXTX Gazamkhan60Încă nu există evaluări
- Manuale Advantage 350Document50 paginiManuale Advantage 350PaulmankeÎncă nu există evaluări
- Memory Manipulation PDFDocument381 paginiMemory Manipulation PDFAlapan Bandyopadhyay100% (2)
- Cardiac MarkersDocument23 paginiCardiac MarkersAsmaaYLÎncă nu există evaluări
- Cancer Fighting StrategiesDocument167 paginiCancer Fighting StrategiesCaptainjillÎncă nu există evaluări
- 165-Article Text-420-1-10-20210830 XXXXDocument6 pagini165-Article Text-420-1-10-20210830 XXXXmokhatikÎncă nu există evaluări
- Hyatt Amritsar showcases Chinese and Thai cuisineDocument1 paginăHyatt Amritsar showcases Chinese and Thai cuisineAnmol MehanÎncă nu există evaluări
- Laparoscopic Appendectomy ReportDocument1 paginăLaparoscopic Appendectomy ReportAjit AJÎncă nu există evaluări
- By hkdsegayau: 通識科 IES 滿分 Sample (樣本)Document10 paginiBy hkdsegayau: 通識科 IES 滿分 Sample (樣本)Chi Shing HoÎncă nu există evaluări
- Nutrition Counseling And Dietary InstructionDocument5 paginiNutrition Counseling And Dietary Instructionnicoleangela ubasroselloÎncă nu există evaluări
- Types and Causes of Medication Errors From A Nurse'S ViewpointDocument3 paginiTypes and Causes of Medication Errors From A Nurse'S ViewpointMaryjoy Gabriellee De La CruzÎncă nu există evaluări
- Diass Prelim ExamDocument9 paginiDiass Prelim ExamLea Luayon Garcia100% (1)
- Cardiovascular ADocument3 paginiCardiovascular AAutumn AllisonÎncă nu există evaluări
- Benefits of Effective Lifting ProgramDocument30 paginiBenefits of Effective Lifting ProgramMoradeke OnasanyaÎncă nu există evaluări
- Colegio de San Gabriel Arcangel, Inc.: READING REPORT in - Complementary and Alternative MedicineDocument2 paginiColegio de San Gabriel Arcangel, Inc.: READING REPORT in - Complementary and Alternative MedicineJonille EchevarriaÎncă nu există evaluări
- Tun Razak Exchange, Retail Plot 1: APPENDIX 5 - Incident Reporting ProcedureDocument5 paginiTun Razak Exchange, Retail Plot 1: APPENDIX 5 - Incident Reporting ProcedureMatthew Mohan PerumalÎncă nu există evaluări
- Eclampsia Nursing Care Plan - Altered Tissue PerfusionDocument2 paginiEclampsia Nursing Care Plan - Altered Tissue PerfusionCyrus De Asis84% (32)
- Water, Sanitation and Hygiene in SchoolsDocument6 paginiWater, Sanitation and Hygiene in SchoolsDux Mercado100% (1)
- The Lord of Lost Heart PDFDocument44 paginiThe Lord of Lost Heart PDFCouteron LaurentÎncă nu există evaluări
- Child Development A Cultural Approach 2nd Edition Arnett Solutions Manual DownloadDocument37 paginiChild Development A Cultural Approach 2nd Edition Arnett Solutions Manual DownloadMichael Pontius100% (24)
- 31.ergonomic Approach of Modification of Seat - EncDocument11 pagini31.ergonomic Approach of Modification of Seat - EncSHASHANK ROHITÎncă nu există evaluări
- What Is Dissociation and What To Do About It?Document2 paginiWhat Is Dissociation and What To Do About It?Tommy ThompsonÎncă nu există evaluări
- Reflection PaperDocument2 paginiReflection Paperapi-355203086Încă nu există evaluări
- Adhaota Vasica (Malabar Nut)Document7 paginiAdhaota Vasica (Malabar Nut)ABHINABA GUPTAÎncă nu există evaluări