S-ar putea să vă placă și
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5795)
- Clinical and Biochemical Footprints of Inherited Metabolic Diseases. II. Metabolic Liver DiseasesDocument5 paginiClinical and Biochemical Footprints of Inherited Metabolic Diseases. II. Metabolic Liver Diseasescarlos1211805752Încă nu există evaluări
- CSF Gradients For Amino Acid NeurotransmittersDocument8 paginiCSF Gradients For Amino Acid Neurotransmitterscarlos1211805752Încă nu există evaluări
- A Case of Congenital Deformity (Dr. Treves' Article On Joseph Merrick)Document12 paginiA Case of Congenital Deformity (Dr. Treves' Article On Joseph Merrick)carlos1211805752Încă nu există evaluări
- Overview of eRA Commons PDFDocument94 paginiOverview of eRA Commons PDFcarlos1211805752Încă nu există evaluări
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
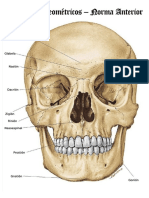
- Puntos Craneometricos CompressDocument13 paginiPuntos Craneometricos CompressLaura PÎncă nu există evaluări
- Mbsimp ScaleDocument3 paginiMbsimp ScaleCamilofonoÎncă nu există evaluări
- Inhs PDFDocument34 paginiInhs PDFKrupa RaoÎncă nu există evaluări
- Toltec Body Healing EbookDocument204 paginiToltec Body Healing EbookDanielle Grace100% (3)
- (Criminalistics 5) Police Polygraphy (Lie-Detection)Document15 pagini(Criminalistics 5) Police Polygraphy (Lie-Detection)Yam P Bustamante100% (2)
- ReceptoretDocument37 paginiReceptoretBlodin ZylfiuÎncă nu există evaluări
- The Peripheral Nervous System Branches Throughout The Body Worksheet1Document2 paginiThe Peripheral Nervous System Branches Throughout The Body Worksheet1Shekeyva SmalleyÎncă nu există evaluări
- Cardio CaseDocument21 paginiCardio CaseJan Rae Y. BattungÎncă nu există evaluări
- School of Out-Of-BodyDocument224 paginiSchool of Out-Of-Bodyrpandit075698100% (2)
- Chapter 24)Document10 paginiChapter 24)Hazel LopezÎncă nu există evaluări
- CarbohydratesDocument10 paginiCarbohydratesshajmalikÎncă nu există evaluări
- Effective Suction-Imporession For Retentive Mand Dentture PDFDocument24 paginiEffective Suction-Imporession For Retentive Mand Dentture PDFFaheemuddin MuhammadÎncă nu există evaluări
- As Biology With Stafford Unit 3 Workbook Answers PDFDocument21 paginiAs Biology With Stafford Unit 3 Workbook Answers PDFFatma ZorluÎncă nu există evaluări
- Anatomical Landmarks of Mandible: DR Hiba HamidDocument29 paginiAnatomical Landmarks of Mandible: DR Hiba HamidDr.Sathyabhama A.VÎncă nu există evaluări
- Pathology of Neurodegenerative DiseasesDocument22 paginiPathology of Neurodegenerative DiseasesKarel GuevaraÎncă nu există evaluări
- Progressions and Methods of Training For YounDocument6 paginiProgressions and Methods of Training For YounIvan NavarroÎncă nu există evaluări
- Cancer Case StudyDocument15 paginiCancer Case StudyRobin HaliliÎncă nu există evaluări
- Learning and Memory 3rd Edition Gluck Test BankDocument26 paginiLearning and Memory 3rd Edition Gluck Test Banklaurenhaynesnjqcdgotay100% (15)
- Cerebrovascular Disease (Emphasis On CVA)Document29 paginiCerebrovascular Disease (Emphasis On CVA)OlynsieMorrisÎncă nu există evaluări
- Zabrina Joana DDocument4 paginiZabrina Joana DEmman BalidoÎncă nu există evaluări
- Human Excretory SystemDocument6 paginiHuman Excretory SystemRahul SinghÎncă nu există evaluări
- Faisalabad Board: Grade 9 Biology 2016 GROUP 1Document11 paginiFaisalabad Board: Grade 9 Biology 2016 GROUP 1DEPUTY DIRECTOR SOCIAL WELFAREÎncă nu există evaluări
- Parenting Children With Reactive Attachment DisorderDocument13 paginiParenting Children With Reactive Attachment Disorderboglous100% (1)
- B1 Quiz 12Document2 paginiB1 Quiz 12giulianotboÎncă nu există evaluări
- Urine Concentration and Dilution Regulation of Extracellular Fluid Osmolarity and Sodium ConcentrationDocument21 paginiUrine Concentration and Dilution Regulation of Extracellular Fluid Osmolarity and Sodium ConcentrationMelissa VillanuevaÎncă nu există evaluări
- Vital SignsDocument3 paginiVital SignsNicole NipasÎncă nu există evaluări
- Bronchiectasis Physiotherapy Assessment Form 2Document4 paginiBronchiectasis Physiotherapy Assessment Form 2jefferyÎncă nu există evaluări
- Complex Anorectal Disorders PDFDocument817 paginiComplex Anorectal Disorders PDFGabriela Balan100% (2)
- 07 Bibliography FormatDocument14 pagini07 Bibliography FormatArjun KafleÎncă nu există evaluări
- Vital Signs 2024Document50 paginiVital Signs 2024gntawfeqÎncă nu există evaluări