S-ar putea să vă placă și
- Mechanism of Action of AriprazoleDocument5 paginiMechanism of Action of AriprazoleSiegfried CristoforÎncă nu există evaluări
- 21-Spinal Cord TractsDocument23 pagini21-Spinal Cord TractsALFAHRUL CAHYADIÎncă nu există evaluări
- 3-Ascending Tracts of Spinal CordDocument28 pagini3-Ascending Tracts of Spinal Cordmuhammad altaf100% (2)
- Somatosensory System: Duu's Topical Diagnosis in NeurologyDocument51 paginiSomatosensory System: Duu's Topical Diagnosis in NeurologySunny AghniÎncă nu există evaluări
- Psychological Science Textbooks: TRIMESTER 3, 2020Document4 paginiPsychological Science Textbooks: TRIMESTER 3, 2020no_if0% (1)
- Spinal Cord LesionsDocument110 paginiSpinal Cord Lesionsgtaha80100% (1)
- Somatosensory PathwaysDocument36 paginiSomatosensory PathwaysAnonymous w3vYureÎncă nu există evaluări
- Lecture 5 p2Document51 paginiLecture 5 p2Ahmed G IsmailÎncă nu există evaluări
- Tracts of The Spinal CordDocument21 paginiTracts of The Spinal Corddr.a.k.gupta6924100% (10)
- TractologyDocument30 paginiTractologyCoskar MethodÎncă nu există evaluări
- Dorsal Column-Medial Lemniscus Pathway - WikipediaDocument27 paginiDorsal Column-Medial Lemniscus Pathway - WikipediaCosplay AccountÎncă nu există evaluări
- 10-Ascending Sensory PATHWAYSDocument12 pagini10-Ascending Sensory PATHWAYSMai OssamaÎncă nu există evaluări
- Sensory and Its DisordersDocument8 paginiSensory and Its Disordersjalan_zÎncă nu există evaluări
- 12 Spinal Cord TractsDocument39 pagini12 Spinal Cord Tractssjs6r8wwv9Încă nu există evaluări
- Somatosensory PathwaysDocument12 paginiSomatosensory PathwaysDimasBanyakRejekiÎncă nu există evaluări
- Neurology TractsDocument7 paginiNeurology TractsMuhammad HaneefÎncă nu există evaluări
- Nerve Tract in Spinal CordDocument30 paginiNerve Tract in Spinal CordirfanzukriÎncă nu există evaluări
- Sensory PathwaysDocument17 paginiSensory PathwaysNabilah Ani SofianÎncă nu există evaluări
- Lecture 5Document22 paginiLecture 5Abdalrhman AhmedÎncă nu există evaluări
- LECTURE On Spinal CordDocument11 paginiLECTURE On Spinal CordElijah KamaniÎncă nu există evaluări
- Anatomia Si Localizare in Boli SpinaleDocument8 paginiAnatomia Si Localizare in Boli SpinaleHalit DianaÎncă nu există evaluări
- Neuroanatomy 1Document65 paginiNeuroanatomy 1maÎncă nu există evaluări
- Sensory and Motor Pathways - DrKatzDocument24 paginiSensory and Motor Pathways - DrKatzWeli PermatasariÎncă nu există evaluări
- Sensory TractsDocument34 paginiSensory TractsMudassar RoomiÎncă nu există evaluări
- Ascending Tracts (Somatosensory Axis)Document94 paginiAscending Tracts (Somatosensory Axis)Gks100% (1)
- Sensory: Sensory (Ascending) Spinal TractsDocument24 paginiSensory: Sensory (Ascending) Spinal Tractsعبد الله الإمامÎncă nu există evaluări
- Posterior Column-Medial Lemniscus PathwayDocument4 paginiPosterior Column-Medial Lemniscus PathwayRachmadina Basyarial BasyaruddinÎncă nu există evaluări
- PY 10.3: Describe and Discuss Sensory TractsDocument34 paginiPY 10.3: Describe and Discuss Sensory TractsJiyaa PatelÎncă nu există evaluări
- Spinal Cord, Ascending PathwayDocument52 paginiSpinal Cord, Ascending PathwayKolawole Adesina KehindeÎncă nu există evaluări
- What Are Spinal Tracts?Document2 paginiWhat Are Spinal Tracts?Ahsan JamilÎncă nu există evaluări
- Spinal Cord: Achmad AminuddinDocument29 paginiSpinal Cord: Achmad AminuddinSurajul AudaÎncă nu există evaluări
- Ascending Spinal Tracts: Dr. Joachim Perera Joachim - Perera@imu - Edu.myDocument26 paginiAscending Spinal Tracts: Dr. Joachim Perera Joachim - Perera@imu - Edu.myZobayer AhmedÎncă nu există evaluări
- Spinal Cord: Navigation SearchDocument16 paginiSpinal Cord: Navigation SearchritzchoiÎncă nu există evaluări
- Anatomi Sistem GerakDocument13 paginiAnatomi Sistem GerakBima AchmadÎncă nu există evaluări
- The Descending Tracts of The Spinal CordDocument2 paginiThe Descending Tracts of The Spinal Cordalmastar officeÎncă nu există evaluări
- CNS and PNSDocument4 paginiCNS and PNSDeepshikhaÎncă nu există evaluări
- Spinal CordDocument48 paginiSpinal CordDr.Saber - Ar - Raffi100% (1)
- Somatic Sensations IDocument23 paginiSomatic Sensations IjolilarmatarÎncă nu există evaluări
- Spinalcord 181202085959Document95 paginiSpinalcord 181202085959gonzalesmailyn85Încă nu există evaluări
- Ascending and Descending Spinal Tracts 2016Document20 paginiAscending and Descending Spinal Tracts 2016Ayodeji LuckyÎncă nu există evaluări
- Sensory System: Dr. Snehal Pallod Guide-Dr - Girish NanotiDocument79 paginiSensory System: Dr. Snehal Pallod Guide-Dr - Girish NanotininoÎncă nu există evaluări
- The Trigeminal NerveDocument19 paginiThe Trigeminal NerveX And ZÎncă nu există evaluări
- Sympathetic Pathway (Thoracolumnar Outflow)Document3 paginiSympathetic Pathway (Thoracolumnar Outflow)NellyWataÎncă nu există evaluări
- Lab. II. Spinal CordDocument7 paginiLab. II. Spinal CordAun CanineÎncă nu există evaluări
- Sensation Kinds and Types of Sensory Disturbance Syndromes of LesionDocument67 paginiSensation Kinds and Types of Sensory Disturbance Syndromes of LesionAhmed Adel SaadÎncă nu există evaluări
- Sensory PathwayDocument31 paginiSensory PathwayAlma BaterinaÎncă nu există evaluări
- Somatosensory System: G.Arvelandze N.MalashkhiaDocument26 paginiSomatosensory System: G.Arvelandze N.MalashkhialuckyÎncă nu există evaluări
- Sistem Saraf Pusat (SSP) Dan Sistem Saraf Tepi (SST)Document37 paginiSistem Saraf Pusat (SSP) Dan Sistem Saraf Tepi (SST)Kris Canter Eka PutraÎncă nu există evaluări
- Guyton Somatic SensesDocument2 paginiGuyton Somatic SensesJamie Ong100% (1)
- Nro NotessafdaadDocument199 paginiNro NotessafdaadMunawwar AwaÎncă nu există evaluări
- Neurology Final QuestionsDocument30 paginiNeurology Final QuestionsyiafkaÎncă nu există evaluări
- Nervous SystemDocument5 paginiNervous SystemAdesinasi Cephas AdewunmiÎncă nu există evaluări
- Neuroscience: Ascending and Descending TracDocument91 paginiNeuroscience: Ascending and Descending Trachazunga rayfordÎncă nu există evaluări
- 11.ascending PathwaysDocument27 pagini11.ascending PathwaysdenekeÎncă nu există evaluări
- The Nervous SystemDocument71 paginiThe Nervous SystemDennis Limosnero MayorÎncă nu există evaluări
- Neuro Oral AnswerDocument113 paginiNeuro Oral AnswerVishva Lalitbhai HirparaÎncă nu există evaluări
- Spinal Tracts: DR - Krishna Madhukar Dept. of Orthopaedics Bharati HospitalDocument65 paginiSpinal Tracts: DR - Krishna Madhukar Dept. of Orthopaedics Bharati HospitalKrishna Madhukar100% (1)
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.De la EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.Încă nu există evaluări
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyDe la EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyÎncă nu există evaluări
- The Secret Life of the Brain: Exploring the Mysteries and Wonders of Our Most Vital OrganDe la EverandThe Secret Life of the Brain: Exploring the Mysteries and Wonders of Our Most Vital OrganÎncă nu există evaluări
- Doa International SeminarDocument2 paginiDoa International Seminardod_nurÎncă nu există evaluări
- EditorsDocument1 paginăEditorsdod_nurÎncă nu există evaluări
- In The First Day of College First Year Medical Students Attend Their First Lecture This MorningDocument1 paginăIn The First Day of College First Year Medical Students Attend Their First Lecture This Morningdod_nurÎncă nu există evaluări
- 1st AnnouncementDocument2 pagini1st Announcementdod_nurÎncă nu există evaluări
- 1-A11.IV-585-S.Kep-UMP-X-2020 - SK HMJDocument3 pagini1-A11.IV-585-S.Kep-UMP-X-2020 - SK HMJdod_nurÎncă nu există evaluări
- Introduction Upper LimbDocument4 paginiIntroduction Upper Limbdod_nurÎncă nu există evaluări
- A 45Document2 paginiA 45dod_nurÎncă nu există evaluări
- Plexus ChoroidalisDocument1 paginăPlexus Choroidalisdod_nurÎncă nu există evaluări
- Encephalo NDocument1 paginăEncephalo Ndod_nurÎncă nu există evaluări
- Skenario Global WarmingDocument5 paginiSkenario Global Warmingdod_nurÎncă nu există evaluări
- AllantoisDocument3 paginiAllantoisdod_nurÎncă nu există evaluări
- UsmleDocument51 paginiUsmleapi-3822318100% (2)
- Ge ItaliaDocument8 paginiGe Italiadod_nurÎncă nu există evaluări
- Soal DigestiveDocument1 paginăSoal Digestivedod_nurÎncă nu există evaluări
- Introduction To Anatomy of MenDocument32 paginiIntroduction To Anatomy of MenRohit KambleÎncă nu există evaluări
- Mortuary FreezerDocument1 paginăMortuary Freezerdod_nurÎncă nu există evaluări
- Cover Laporan Pengabdian MasyarakatDocument2 paginiCover Laporan Pengabdian Masyarakatdod_nurÎncă nu există evaluări
- Around The Time of The Twelfth Congress Nomina AnatomicaDocument2 paginiAround The Time of The Twelfth Congress Nomina Anatomicadod_nurÎncă nu există evaluări
- DigestifDocument3 paginiDigestifdod_nurÎncă nu există evaluări
- Wave Lab InstalDocument1 paginăWave Lab Instaldod_nurÎncă nu există evaluări
- The Hominids: © 2008 Paul BillietDocument19 paginiThe Hominids: © 2008 Paul Billietdod_nurÎncă nu există evaluări
- Data Shet SynaptophysinsDocument4 paginiData Shet Synaptophysinsdod_nurÎncă nu există evaluări
- 19 CadaversDocument14 pagini19 CadaversPatrick ArleyÎncă nu există evaluări
- 1181Document19 pagini1181Anne KawÎncă nu există evaluări
- Dev 2Document12 paginiDev 2dod_nurÎncă nu există evaluări
- Dev 2Document12 paginiDev 2dod_nurÎncă nu există evaluări
- 242 Full PDFDocument16 pagini242 Full PDFdod_nurÎncă nu există evaluări
- Bio DevelopmentalDocument9 paginiBio Developmentaldod_nurÎncă nu există evaluări
- Journal Developmental BioDocument11 paginiJournal Developmental Biodod_nurÎncă nu există evaluări
- DevelopmentDocument8 paginiDevelopmentdod_nurÎncă nu există evaluări
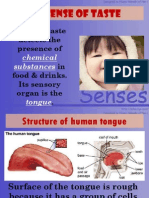
- 1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheDocument6 pagini1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheMissz WazeeÎncă nu există evaluări
- Common Types of Tests On Human CognitionDocument9 paginiCommon Types of Tests On Human CognitionNesrin MemedÎncă nu există evaluări
- PDEV 111 Answer 2nd QuarterDocument30 paginiPDEV 111 Answer 2nd QuarterChan-Chan Matamorosa50% (2)
- Assignment: Riphah International University (Faisalabad Campus)Document5 paginiAssignment: Riphah International University (Faisalabad Campus)Muhammad Uzair GujjarÎncă nu există evaluări
- Artificial Intelligence - Curriculum PDFDocument9 paginiArtificial Intelligence - Curriculum PDFNitika KapoorÎncă nu există evaluări
- Contoh - Laporan - Bab 2Document3 paginiContoh - Laporan - Bab 2Ayu komala sariÎncă nu există evaluări
- ParamnesiaDocument3 paginiParamnesiaThe LullabyÎncă nu există evaluări
- Learning Theory of Cognitivism and Its Implications On Students Learning - 1622555691Document5 paginiLearning Theory of Cognitivism and Its Implications On Students Learning - 1622555691Azurawati WokZakiÎncă nu există evaluări
- Richard Feynman On DreamsDocument5 paginiRichard Feynman On DreamsDrBertram ForerÎncă nu există evaluări
- Problem Solving Graphic OrganizerDocument5 paginiProblem Solving Graphic Organizerapi-458722870Încă nu există evaluări
- 1-External Features of Spinal CordDocument24 pagini1-External Features of Spinal Cordyaram3512Încă nu există evaluări
- Pple Assessment 2 Reflection - Philosophy of Classroom ManagementDocument16 paginiPple Assessment 2 Reflection - Philosophy of Classroom Managementapi-331237685Încă nu există evaluări
- Types of Learning Style ModelsDocument5 paginiTypes of Learning Style ModelsJaypee Bitun EstalÎncă nu există evaluări
- 25 OT Frames of Reference in Brief - Ram, APDocument23 pagini25 OT Frames of Reference in Brief - Ram, APS.JAIVIGNESH OTÎncă nu există evaluări
- Power of Energy HealingDocument4 paginiPower of Energy HealingBeyond ThoughtsÎncă nu există evaluări
- Reflection Ni Joana Rose QueddengDocument3 paginiReflection Ni Joana Rose Queddengjhon ryanÎncă nu există evaluări
- Clerking TemplateDocument4 paginiClerking TemplateNur Atiqah Mohd Azli100% (1)
- Research StudyDocument12 paginiResearch Studysniperhunter881Încă nu există evaluări
- Efferent Visual System (Ocular Motor Pathways)Document38 paginiEfferent Visual System (Ocular Motor Pathways)riskhapangestika100% (1)
- Demonet 2005 Neurofisiologia Lenguaje IMPRESODocument47 paginiDemonet 2005 Neurofisiologia Lenguaje IMPRESOOscar M. Cassiane CamachoÎncă nu există evaluări
- Erotictransference: Alberto StefanaDocument9 paginiErotictransference: Alberto StefanaRozita RoziniÎncă nu există evaluări
- Final NeuronotesDocument16 paginiFinal Neuronotesapi-479939114Încă nu există evaluări
- Ap Psych Exam Review SheetDocument4 paginiAp Psych Exam Review Sheetapi-31355470Încă nu există evaluări
- Effect of Vergence Adaptation and Positive Fusional Vergence Training On Oculomotor ParametersDocument121 paginiEffect of Vergence Adaptation and Positive Fusional Vergence Training On Oculomotor ParametersGian Carlo Giusto BurboaÎncă nu există evaluări
- Teaching Materials: Grade 1Document1 paginăTeaching Materials: Grade 1bayviewÎncă nu există evaluări
- Nervous System of FishDocument4 paginiNervous System of Fishtalam_06100% (10)
- Reading Sample Test 4 Answer Key Part BCDocument2 paginiReading Sample Test 4 Answer Key Part BCNanda NandaÎncă nu există evaluări
- A Term Presented To The Faculty of The College of Teacher Education University of The CordillerasDocument7 paginiA Term Presented To The Faculty of The College of Teacher Education University of The CordillerasErna SalazarÎncă nu există evaluări