S-ar putea să vă placă și
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (121)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Bush y FrancisDocument2 paginiBush y FrancisCarol Artigas100% (1)
- CHCCCS015 Student Assessment Booklet Is (ID 97088) - FinalDocument33 paginiCHCCCS015 Student Assessment Booklet Is (ID 97088) - FinalESRÎncă nu există evaluări
- Cytogenetics - Prelim TransesDocument15 paginiCytogenetics - Prelim TransesLOUISSE ANNE MONIQUE L. CAYLOÎncă nu există evaluări
- Human Body Systems: Study GuideDocument11 paginiHuman Body Systems: Study Guideapi-242114183Încă nu există evaluări
- 2 - Cannabis Intoxication Case SeriesDocument8 pagini2 - Cannabis Intoxication Case SeriesPhelipe PessanhaÎncă nu există evaluări
- Conduction Blocks in Acute Myocardial Infarction: A Prospective StudyDocument6 paginiConduction Blocks in Acute Myocardial Infarction: A Prospective StudyJack JacksonÎncă nu există evaluări
- Enfermedades Productos o AfeccionesDocument9 paginiEnfermedades Productos o AfeccionesHumberto Colque LlaveraÎncă nu există evaluări
- 8mkebook PDFDocument300 pagini8mkebook PDFVanja Vujovic100% (2)
- New Study in Candida InfectionsDocument3 paginiNew Study in Candida InfectionsHenz FreemanÎncă nu există evaluări
- Towards A Sociology of Health Discourse in AfricaDocument172 paginiTowards A Sociology of Health Discourse in AfricaDavid Polowiski100% (2)
- Maxime Gorky and KingstonDocument433 paginiMaxime Gorky and KingstonBasil FletcherÎncă nu există evaluări
- Menstrual Blood Derived Stem Cells and Their Scope in Regenerative Medicine A Review ArticleDocument6 paginiMenstrual Blood Derived Stem Cells and Their Scope in Regenerative Medicine A Review ArticleInternational Journal of Innovative Science and Research TechnologyÎncă nu există evaluări
- Dr. Sudarto Sppd's FileDocument22 paginiDr. Sudarto Sppd's FileVindy CesarianaÎncă nu există evaluări
- HED 2007 Cellular Molecular, Microbiology & GeneticsDocument16 paginiHED 2007 Cellular Molecular, Microbiology & Geneticsharyshan100% (1)
- The Nadi Vigyan by DR - Sharda Mishra MD (Proff. in Jabalpur Ayurved College)Document5 paginiThe Nadi Vigyan by DR - Sharda Mishra MD (Proff. in Jabalpur Ayurved College)Vivek PandeyÎncă nu există evaluări
- The Human Magnet Syndrome Why We Love PeDocument8 paginiThe Human Magnet Syndrome Why We Love Peapi-316440587Încă nu există evaluări
- Cell Types Animal Tissue ReviewerDocument6 paginiCell Types Animal Tissue ReviewerShane LaluÎncă nu există evaluări
- Aldrine Ilustricimo VS Nyk Fil Sjip Management IncDocument11 paginiAldrine Ilustricimo VS Nyk Fil Sjip Management Inckristel jane caldozaÎncă nu există evaluări
- Tendonitis Guide - Causes, Symptoms and Treatment OptionsDocument6 paginiTendonitis Guide - Causes, Symptoms and Treatment OptionsSylvia GraceÎncă nu există evaluări
- Beauty Hankering For Memory: Marne L. KilatesDocument9 paginiBeauty Hankering For Memory: Marne L. KilatesKuya ReaGoÎncă nu există evaluări
- Msds ChloroformDocument9 paginiMsds ChloroformAhmad ArisandiÎncă nu există evaluări
- Class IX - Worksheet 4 (Comprehension & Writing Skill)Document6 paginiClass IX - Worksheet 4 (Comprehension & Writing Skill)Anis FathimaÎncă nu există evaluări
- Sample Questionnaire For Understanding The Self: For Private Use OnlyDocument6 paginiSample Questionnaire For Understanding The Self: For Private Use Onlyirene ilustrisimo100% (1)
- What Is Malnutrition?: WastingDocument6 paginiWhat Is Malnutrition?: WastingĐoan VõÎncă nu există evaluări
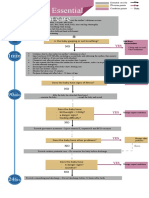
- Algorithm: Essential Newborn Care: Is The Baby Gasping or Not Breathing?Document1 paginăAlgorithm: Essential Newborn Care: Is The Baby Gasping or Not Breathing?Dela cruz KimberlyÎncă nu există evaluări
- Journals With No Article Processing ChargesDocument14 paginiJournals With No Article Processing ChargesNurul Jami Bakti JayaÎncă nu există evaluări
- A Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaDocument8 paginiA Comparative Clinical Evaluation of Eranda Taila Matra Basti With Combination of Mefenamic Acid and Dicyclomine in The Management of Udavartini W.S.R To Primary DysmenorrheaEditor IJTSRDÎncă nu există evaluări
- A Manual 20199 CompleetDocument201 paginiA Manual 20199 CompleetJanPerezÎncă nu există evaluări
- LAN Party Skate Park by Shane Jesse ChristmassDocument91 paginiLAN Party Skate Park by Shane Jesse ChristmassPatrick TrottiÎncă nu există evaluări
- Dula-Tungkulin o Gampanin NG ProduksyonDocument12 paginiDula-Tungkulin o Gampanin NG ProduksyonBernadette DuranÎncă nu există evaluări