S-ar putea să vă placă și
- Understanding CHF and Monitoring Fluid StatusDocument4 paginiUnderstanding CHF and Monitoring Fluid StatusWeng RamojalÎncă nu există evaluări
- ... & More: ECG ChallengeDocument2 pagini... & More: ECG ChallengehameunjungÎncă nu există evaluări
- Fatime Sanogo Vsim Steps - HTMLDocument6 paginiFatime Sanogo Vsim Steps - HTMLJhunnieEy ReyesÎncă nu există evaluări
- Vital Signs Reference Chart 1.2 - 1 PDFDocument1 paginăVital Signs Reference Chart 1.2 - 1 PDFSerious Leo100% (1)
- A Presentation On Kit KatDocument9 paginiA Presentation On Kit KatRaju DoleyÎncă nu există evaluări
- Tank Cleaning ConsiderationsDocument1 paginăTank Cleaning ConsiderationsAdele PollardÎncă nu există evaluări
- Vascular Responses to PathogensDe la EverandVascular Responses to PathogensFelicity N.E. GavinsÎncă nu există evaluări
- Mesenteric Ischemia in Dilated CardiomyopathyDocument30 paginiMesenteric Ischemia in Dilated CardiomyopathyJo AnneÎncă nu există evaluări
- Peds Lecture 3Document19 paginiPeds Lecture 3Suraj Mukatira100% (1)
- Quiz 2 Study Guide - SHOCK!!! 25 Questions READ BOOK!!! Know Charts Esp. Stages of Shock ChartDocument1 paginăQuiz 2 Study Guide - SHOCK!!! 25 Questions READ BOOK!!! Know Charts Esp. Stages of Shock Chartjenn1722Încă nu există evaluări
- Assignmen 1 AnswersDocument3 paginiAssignmen 1 AnswersAlasl BobyÎncă nu există evaluări
- IV Lab Practice Problems Answers SP 2013 PALDocument2 paginiIV Lab Practice Problems Answers SP 2013 PALjmraczynskiÎncă nu există evaluări
- A. Cardiac Failure: Biologic CrisisDocument11 paginiA. Cardiac Failure: Biologic CrisisJillian CaumbanÎncă nu există evaluări
- Gastro MnemonicsDocument8 paginiGastro MnemonicsRufina SoomroÎncă nu există evaluări
- Medical-Surgical Nursing Review: 315 Key ConceptsDocument15 paginiMedical-Surgical Nursing Review: 315 Key ConceptsJap Avila BernasÎncă nu există evaluări
- Concept Map Et Al 11-04-15Document7 paginiConcept Map Et Al 11-04-15api-353656227Încă nu există evaluări
- What Is Respiratory Alkalosis?Document3 paginiWhat Is Respiratory Alkalosis?Lorenn AdarnaÎncă nu există evaluări
- Medsurg Test 4 ReviewDocument11 paginiMedsurg Test 4 ReviewTori RolandÎncă nu există evaluări
- Congestive Heart Failure OverviewDocument12 paginiCongestive Heart Failure OverviewkazelleÎncă nu există evaluări
- Anatomy, Diagnostics and Nursing Care of Pancreatitis and Gallbladder DisordersDocument21 paginiAnatomy, Diagnostics and Nursing Care of Pancreatitis and Gallbladder DisordersKatrinaJimenezÎncă nu există evaluări
- High Blood Pressure GuideDocument4 paginiHigh Blood Pressure GuideKhaira CharmagneÎncă nu există evaluări
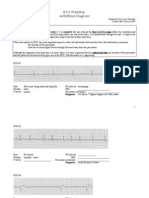
- ECG Workshop 2006Document15 paginiECG Workshop 2006Eggi ErlanggaÎncă nu există evaluări
- HESI LPN ADN ENTRANCE EXAM MOBILITY EXAMS A GRADED 100 VERIFIED - pdf-1-33Document33 paginiHESI LPN ADN ENTRANCE EXAM MOBILITY EXAMS A GRADED 100 VERIFIED - pdf-1-33erick kanyiÎncă nu există evaluări
- Hematology 101: Hematology/Oncology PG 1 of 11Document12 paginiHematology 101: Hematology/Oncology PG 1 of 11esther100% (1)
- MS Final 46 Blood or Lymphatic DisorderDocument4 paginiMS Final 46 Blood or Lymphatic DisorderZachary T Hall0% (1)
- Grand Rounds - Pulmonary EmbolismDocument25 paginiGrand Rounds - Pulmonary EmbolismAdrian CastroÎncă nu există evaluări
- Main drug therapy for ulcerative colitis is sulfa for one yearDocument4 paginiMain drug therapy for ulcerative colitis is sulfa for one yearcathyÎncă nu există evaluări
- Tocolytic Agents and Anticonvulsants for Preterm LabourDocument95 paginiTocolytic Agents and Anticonvulsants for Preterm LabourPriya jÎncă nu există evaluări
- Congestive Heart Failure LectureDocument102 paginiCongestive Heart Failure Lectureluke100% (1)
- Conduction System: Rhythm Identification and TreatmentDocument12 paginiConduction System: Rhythm Identification and Treatmenthops23Încă nu există evaluări
- Case Study: Congestive Heart FailureDocument7 paginiCase Study: Congestive Heart FailureXI-E / 21 / MARY TRIANAÎncă nu există evaluări
- Managing Angina: AHA Guidelines on Diagnosis and TreatmentDocument96 paginiManaging Angina: AHA Guidelines on Diagnosis and TreatmentJaymica Laggui DacquilÎncă nu există evaluări
- PedsnotesDocument18 paginiPedsnoteskp13oyÎncă nu există evaluări
- 224 Cardio Exam II - 2Document29 pagini224 Cardio Exam II - 2fiya33Încă nu există evaluări
- Hypertension PDFDocument19 paginiHypertension PDFVicta RyzaÎncă nu există evaluări
- Syncope - Approach To The Patient Dynamed 2020 PDFDocument76 paginiSyncope - Approach To The Patient Dynamed 2020 PDFHeriberto Moreno HernandezÎncă nu există evaluări
- Care PlanDocument11 paginiCare PlanAlyssa CardinalÎncă nu există evaluări
- Major Hormone Glands and Their FunctionsDocument5 paginiMajor Hormone Glands and Their FunctionsSTEFFI GABRIELLE GOLEZÎncă nu există evaluări
- Shock Types 141009102815 Conversion Gate01Document41 paginiShock Types 141009102815 Conversion Gate01Samjaisheel SamsonÎncă nu există evaluări
- Paeds Handbook Class2021 1568487638Document30 paginiPaeds Handbook Class2021 1568487638kafosidÎncă nu există evaluări
- ShockDocument21 paginiShockMin-Joo Esther ParkÎncă nu există evaluări
- Congenital Heart DiseaseDocument45 paginiCongenital Heart DiseaseBrandedlovers OnlineshopÎncă nu există evaluări
- BluePrint 5th QuestionsDocument25 paginiBluePrint 5th QuestionsAbuFreihÎncă nu există evaluări
- Addison and Cushing DMDocument22 paginiAddison and Cushing DManon_391984943Încă nu există evaluări
- PBL PgamboaDocument6 paginiPBL PgamboaLeanne Princess GamboaÎncă nu există evaluări
- 5 Digestive IDocument11 pagini5 Digestive IMoataz TrabehÎncă nu există evaluări
- Congestive Heart FailureDocument19 paginiCongestive Heart FailureIlavenil PanduranganÎncă nu există evaluări
- CorrectsDocument46 paginiCorrectsnursing_hallÎncă nu există evaluări
- Management of Severe Hypertension, Hypertension in Special ConditionDocument43 paginiManagement of Severe Hypertension, Hypertension in Special Conditionabhandlung100% (3)
- Neuro, GI, GU, MSK midterm and final reviewDocument3 paginiNeuro, GI, GU, MSK midterm and final reviewjenn1722Încă nu există evaluări
- Cad PPTDocument81 paginiCad PPTvaishnaviÎncă nu există evaluări
- Chapter 25heart Failure and Inflammatory DysfunctionDocument10 paginiChapter 25heart Failure and Inflammatory DysfunctionNurse UtopiaÎncă nu există evaluări
- Quiz With AnswersDocument8 paginiQuiz With Answersval284yÎncă nu există evaluări
- Potassium and Sodium Level Imbalances: Hyperkalemia, Hypokalemia, Hypernatremia, HyponatremiaDocument5 paginiPotassium and Sodium Level Imbalances: Hyperkalemia, Hypokalemia, Hypernatremia, HyponatremiaChariza Trompeta100% (1)
- Study Guide For Med Surg 1Document15 paginiStudy Guide For Med Surg 1desireemaenugentÎncă nu există evaluări
- Respiratory meds guide for new nursesDocument2 paginiRespiratory meds guide for new nursesmlbrown8Încă nu există evaluări
- ILOCOS TRAINING AND REGIONAL MEDICAL CENTER JUNIOR INTERNS’ EXIT EXAMINATIONDocument6 paginiILOCOS TRAINING AND REGIONAL MEDICAL CENTER JUNIOR INTERNS’ EXIT EXAMINATIONKai SibayanÎncă nu există evaluări
- HTNDocument15 paginiHTNNatukunda DianahÎncă nu există evaluări
- Necrotizing Otitis 2022Document20 paginiNecrotizing Otitis 2022asmashÎncă nu există evaluări
- HTNDocument54 paginiHTNapi-232466940Încă nu există evaluări
- Myocardial Infarction QuestionsDocument10 paginiMyocardial Infarction Questionsdsaitta108Încă nu există evaluări
- A Simple Guide to Abdominal Aortic Aneurysm, Diagnosis, Treatment and Related ConditionsDe la EverandA Simple Guide to Abdominal Aortic Aneurysm, Diagnosis, Treatment and Related ConditionsÎncă nu există evaluări
- A Simple Guide to Hypovolemia, Diagnosis, Treatment and Related ConditionsDe la EverandA Simple Guide to Hypovolemia, Diagnosis, Treatment and Related ConditionsÎncă nu există evaluări
- 1753 2000 2 13Document7 pagini1753 2000 2 13BimaIndraÎncă nu există evaluări
- Lembar KonsultasiDocument16 paginiLembar KonsultasihskaialsbsusjÎncă nu există evaluări
- ArrestDocument9 paginiArrestBimaIndraÎncă nu există evaluări
- Jurnal Shintia PDFDocument13 paginiJurnal Shintia PDFBimaIndraÎncă nu există evaluări
- Complications Phototherapy Hyperbilirubinaemia: NeonatalDocument3 paginiComplications Phototherapy Hyperbilirubinaemia: NeonatalBimaIndraÎncă nu există evaluări
- WHO PPHDocument62 paginiWHO PPHAchyut SharmaÎncă nu există evaluări
- Patient room assignmentsDocument5 paginiPatient room assignmentsBimaIndraÎncă nu există evaluări
- Urinary Tract Infections Guide for Diagnosis and TreatmentDocument22 paginiUrinary Tract Infections Guide for Diagnosis and TreatmentpupuayuwandiraÎncă nu există evaluări
- Patient room assignmentsDocument5 paginiPatient room assignmentsBimaIndraÎncă nu există evaluări
- GTG 50 Umbilicalcordprolapse 2014Document19 paginiGTG 50 Umbilicalcordprolapse 2014garfield1Încă nu există evaluări
- Patient room assignmentsDocument5 paginiPatient room assignmentsBimaIndraÎncă nu există evaluări
- Congenital Cardiovascular Defects: Statistical Fact Sheet 2016 UpdateDocument2 paginiCongenital Cardiovascular Defects: Statistical Fact Sheet 2016 UpdateBimaIndraÎncă nu există evaluări
- Patient room assignmentsDocument5 paginiPatient room assignmentsBimaIndraÎncă nu există evaluări
- 1Document13 pagini1alviantiauliaÎncă nu există evaluări
- Cord ProlapseDocument15 paginiCord Prolapsexxdrivexx67% (3)
- 3Document12 pagini3alviantiauliaÎncă nu există evaluări
- Arrhythmias Consist of Cardiac Depolarizations That Deviate From The Above Description in One or More AspectsDocument4 paginiArrhythmias Consist of Cardiac Depolarizations That Deviate From The Above Description in One or More AspectsBimaIndraÎncă nu există evaluări
- Pediatricreferencecard 04Document2 paginiPediatricreferencecard 04yesumovsÎncă nu există evaluări
- NeurofibromatosisDocument1 paginăNeurofibromatosisBimaIndraÎncă nu există evaluări
- Listening PutihDocument2 paginiListening PutihAhmad Zulkarnain ArikoÎncă nu există evaluări
- Tuli Sensorineural / Sensorineural Hearing LossDocument25 paginiTuli Sensorineural / Sensorineural Hearing Lossdika koswara80% (5)
- NeurofibromatosisDocument6 paginiNeurofibromatosisBimaIndraÎncă nu există evaluări
- Krisis TiroidDocument24 paginiKrisis TiroidBimaIndraÎncă nu există evaluări
- HalothaneDocument7 paginiHalothaneBimaIndraÎncă nu există evaluări
- INTERNAL CAPSULE TUTORIALDocument24 paginiINTERNAL CAPSULE TUTORIALBimaIndraÎncă nu există evaluări
- HalothaneDocument7 paginiHalothaneBimaIndraÎncă nu există evaluări
- ClopidogrelDocument20 paginiClopidogrelBimaIndraÎncă nu există evaluări
- Depok: Departemen Farmasi FMIPA-UI, Universitas IndonesiaDocument1 paginăDepok: Departemen Farmasi FMIPA-UI, Universitas IndonesiaBimaIndraÎncă nu există evaluări
- Comparison Between China and Sri-Lanka GDPDocument6 paginiComparison Between China and Sri-Lanka GDPcracking khalifÎncă nu există evaluări
- Ketamine Drug Study for AnesthesiaDocument1 paginăKetamine Drug Study for AnesthesiaPRINCESS MARIZHAR OMARÎncă nu există evaluări
- Checkpoint 2015 Paper 1Document6 paginiCheckpoint 2015 Paper 1Aries SaepanÎncă nu există evaluări
- HLF Sharing Fitness Business CooperationDocument17 paginiHLF Sharing Fitness Business Cooperationbhardwajharsh2706Încă nu există evaluări
- ZCT ZCT ZCT ZCT: 40S 60S 80S 120S 210SDocument1 paginăZCT ZCT ZCT ZCT: 40S 60S 80S 120S 210SWilliam TanÎncă nu există evaluări
- Experimental Animals Vol1Document209 paginiExperimental Animals Vol1JohnÎncă nu există evaluări
- Non-Directive Counselling RogersDocument5 paginiNon-Directive Counselling RogersSanjeev ChaudharyÎncă nu există evaluări
- MANUAL Health O Meter Scale 800KLDocument2 paginiMANUAL Health O Meter Scale 800KLElkin MaldonadoÎncă nu există evaluări
- Review of LitetaureDocument8 paginiReview of LitetaureeswariÎncă nu există evaluări
- Register Environmental ImpactsDocument7 paginiRegister Environmental ImpactsArmand LiviuÎncă nu există evaluări
- Report Information From Proquest: February 24 2014 06:18Document19 paginiReport Information From Proquest: February 24 2014 06:18Emma Elena StroeÎncă nu există evaluări
- One Stop English - Cornish Village - Pre-IntermediateDocument5 paginiOne Stop English - Cornish Village - Pre-Intermediatec_a_tabetÎncă nu există evaluări
- GarrettDocument2 paginiGarrettAndrey MarviantoÎncă nu există evaluări
- Causes & Prevention of Coronary Artery DiseaseDocument41 paginiCauses & Prevention of Coronary Artery DiseaseeenagpurcongÎncă nu există evaluări
- Technical PaperDocument10 paginiTechnical Paperalfosoa5505Încă nu există evaluări
- Step by Step To The Perfect PedicureDocument6 paginiStep by Step To The Perfect PedicurepinkyÎncă nu există evaluări
- wk4 Stdconferencereflection ElmoreajaDocument4 paginiwk4 Stdconferencereflection Elmoreajaapi-316378224Încă nu există evaluări
- Everything You Want To Know in ChennaiDocument428 paginiEverything You Want To Know in ChennaiBalamuruganÎncă nu există evaluări
- Liu030 Nepal Bans Solo Mountain ClimbersDocument2 paginiLiu030 Nepal Bans Solo Mountain Climberssanti.miranda.parrillaÎncă nu există evaluări
- Hawthorne SurveyDocument2 paginiHawthorne Surveyapi-256186318Încă nu există evaluări
- Cloudsoc For Amazon Web Services Solution Overview enDocument6 paginiCloudsoc For Amazon Web Services Solution Overview enmanishÎncă nu există evaluări
- WILLIEEMS TIBLANI - NURS10 Student Copy Module 15 Part1Document32 paginiWILLIEEMS TIBLANI - NURS10 Student Copy Module 15 Part1Toyour EternityÎncă nu există evaluări
- HPAD 201 Sat Days 2 and 3Document11 paginiHPAD 201 Sat Days 2 and 3Arianne A ZamoraÎncă nu există evaluări
- ApiDocument4 paginiApiMahmoud Alwasif100% (1)
- NPD High Level Status: Concept Development FeasibilityDocument22 paginiNPD High Level Status: Concept Development FeasibilityRaviÎncă nu există evaluări
- EVA and MVADocument19 paginiEVA and MVATanveer Ahmad100% (1)
- Forensic Science Project Group B5518Document5 paginiForensic Science Project Group B5518Anchit JassalÎncă nu există evaluări
- MBBS Final Part-I (Winter Session) Time Table (Jan 2023)Document1 paginăMBBS Final Part-I (Winter Session) Time Table (Jan 2023)crystal mindÎncă nu există evaluări