S-ar putea să vă placă și
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Graft RejectionDocument8 paginiGraft Rejectionasmaa100% (1)
- CL HNDBDS 08 22 (Literature Review)Document18 paginiCL HNDBDS 08 22 (Literature Review)Biseka De SilvaÎncă nu există evaluări
- Cladogram CH 19 Part ADocument3 paginiCladogram CH 19 Part AJasmine lee100% (1)
- Primary Lymphoid Organs Generate Immune CellsDocument10 paginiPrimary Lymphoid Organs Generate Immune CellsKarthigaÎncă nu există evaluări
- Molecular Biology: Louise Kamona Kabwe 1616713: 1001839253:::: / 784199656856434: 100821ASTJ0035Document1 paginăMolecular Biology: Louise Kamona Kabwe 1616713: 1001839253:::: / 784199656856434: 100821ASTJ0035Louise KamonaÎncă nu există evaluări
- Cell Transport Practice TestDocument4 paginiCell Transport Practice TestLavander Blush100% (2)
- Test I. Blood Puzzle: Find The Words Listed Below in The Word SearchDocument2 paginiTest I. Blood Puzzle: Find The Words Listed Below in The Word SearchPilarieBernalÎncă nu există evaluări
- A New Evolutionary Law of Constant Extinction RatesDocument30 paginiA New Evolutionary Law of Constant Extinction RatesMarcos Vinicius Monteiro100% (1)
- Topic - 8Document29 paginiTopic - 8Cynthia GÎncă nu există evaluări
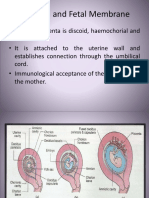
- Placenta and Fetal Membrane-6614Document45 paginiPlacenta and Fetal Membrane-6614Incredible DivineÎncă nu există evaluări
- Bacteriological Analysis of Water: MPN Technique for Coliform CountDocument9 paginiBacteriological Analysis of Water: MPN Technique for Coliform CountGaurav MudaduÎncă nu există evaluări
- UGCNET Paper2 AnthropologyDocument56 paginiUGCNET Paper2 AnthropologyAryanÎncă nu există evaluări
- Electrophysiological Recording Techniques PDFDocument7 paginiElectrophysiological Recording Techniques PDFAndrei TatomirÎncă nu există evaluări
- Function of RBCDocument9 paginiFunction of RBCAthea MelosantosÎncă nu există evaluări
- 1 - Brief Biography of Charles Darwin (1809 - 1882)Document22 pagini1 - Brief Biography of Charles Darwin (1809 - 1882)vladÎncă nu există evaluări
- Materi 1 KLS Xi PersonalDocument4 paginiMateri 1 KLS Xi Personalakusopo JalÎncă nu există evaluări
- Effects of High Temperature On Nodulation and Nitrogen Fixation byDocument8 paginiEffects of High Temperature On Nodulation and Nitrogen Fixation byFábio Luís MostassoÎncă nu există evaluări
- 2110 IAL Subject Grade BoundariesDocument6 pagini2110 IAL Subject Grade BoundariesAshijÎncă nu există evaluări
- Gong Et Al. - Two New Garra Species From Yarlung TsangpoDocument26 paginiGong Et Al. - Two New Garra Species From Yarlung TsangpoSubhadra LaimayumÎncă nu există evaluări
- The Kingdom Protoctista (Protista)Document4 paginiThe Kingdom Protoctista (Protista)MSÎncă nu există evaluări
- L. plantarum 299v Reduces Inflammation and Normalizes Motility in IBS Mouse ModelDocument6 paginiL. plantarum 299v Reduces Inflammation and Normalizes Motility in IBS Mouse ModelMarin GregurinčićÎncă nu există evaluări
- (Studies in Behavioural Adaptation) Robin I. M. Dunbar (Auth.) - Primate Social SystemDocument382 pagini(Studies in Behavioural Adaptation) Robin I. M. Dunbar (Auth.) - Primate Social SystemSourav Kapri100% (1)
- Drug DesignDocument38 paginiDrug DesignPhArMaCyGrAdUaTeSÎncă nu există evaluări
- PlantsDocument84 paginiPlantsYanyan PaydaÎncă nu există evaluări
- Snyder - Science and Psychiatry - Groundbreaking Discoveries in Molecular Neuroscience PDFDocument514 paginiSnyder - Science and Psychiatry - Groundbreaking Discoveries in Molecular Neuroscience PDFuterw100% (1)
- 1730 3631 1 PBDocument10 pagini1730 3631 1 PB19-2024 Ananda Hera UtamaÎncă nu există evaluări
- Recent Advances in Periodontal Diagnosis-SoniaDocument143 paginiRecent Advances in Periodontal Diagnosis-SoniaPuneet SachdevaÎncă nu există evaluări
- Bacterial ReproductionDocument12 paginiBacterial Reproductionchann.maahiÎncă nu există evaluări
- Yamada, Yoshio Et Al. 1966Document24 paginiYamada, Yoshio Et Al. 1966akumakujaÎncă nu există evaluări
- Understanding Apicomplexan ParasitesDocument51 paginiUnderstanding Apicomplexan ParasitesruthÎncă nu există evaluări