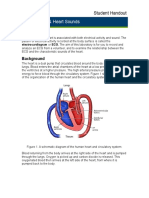
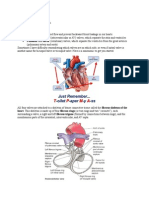
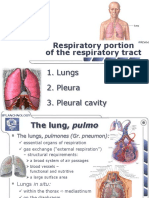
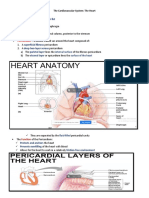
S-ar putea să vă placă și
- 1st Lec On Heart Physiology by Dr. RoomiDocument21 pagini1st Lec On Heart Physiology by Dr. RoomiMudassar RoomiÎncă nu există evaluări
- Lung Metabolism: Proteolysis and Antioproteolysis Biochemical Pharmacology Handling of Bioactive SubstancesDe la EverandLung Metabolism: Proteolysis and Antioproteolysis Biochemical Pharmacology Handling of Bioactive SubstancesAlain JunodÎncă nu există evaluări
- The Artificial Heart: A Design Example: BIOE 1000 October 18, 2001Document17 paginiThe Artificial Heart: A Design Example: BIOE 1000 October 18, 2001Paspulati Leelaram100% (1)
- Unit 1 - Cardiac Care UnitsDocument29 paginiUnit 1 - Cardiac Care UnitssitalekshmiÎncă nu există evaluări
- NMR to MRI: Impact of the Second World War on Studies of Magnetic ResonanceDe la EverandNMR to MRI: Impact of the Second World War on Studies of Magnetic ResonanceÎncă nu există evaluări
- Cardiac PhysiologyDocument102 paginiCardiac PhysiologyPeter Larsen100% (1)
- Cardiac Output: Dr. AthulyaDocument29 paginiCardiac Output: Dr. AthulyaamrendraÎncă nu există evaluări
- Fisiologia CardiovascularDocument94 paginiFisiologia CardiovascularDenisse FimbresÎncă nu există evaluări
- Introduction To Biopotential Measurement and Excitation: (Bioelectromagnetism - Jakko Malmivuo, Robert Plonsey)Document23 paginiIntroduction To Biopotential Measurement and Excitation: (Bioelectromagnetism - Jakko Malmivuo, Robert Plonsey)Lynn WankharÎncă nu există evaluări
- Participate in Board Questions: Ascexam / ReasceDocument105 paginiParticipate in Board Questions: Ascexam / ReasceSharma RomeoÎncă nu există evaluări
- Simple Office SpirometryDocument42 paginiSimple Office SpirometryRoberto Merza III100% (4)
- ECG & Heart Sounds: Student HandoutDocument7 paginiECG & Heart Sounds: Student HandoutROHITÎncă nu există evaluări
- Chapter 9 Cardiac MuscleDocument5 paginiChapter 9 Cardiac MuscleYheng GaosaiiÎncă nu există evaluări
- Nerve TissueDocument148 paginiNerve TissuePatriciaMariskaKrisnawatiSenjaya100% (1)
- Hemodynamics: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeDocument31 paginiHemodynamics: DR Axelle Saverettiar Mbbs Physiology Department SSR Medical CollegeÑäd ÉèmÎncă nu există evaluări
- Copy1 TOPANAT EXAM MCQ Nee RevisedDocument125 paginiCopy1 TOPANAT EXAM MCQ Nee Revisedkapil pancholiÎncă nu există evaluări
- Blood VesselsDocument7 paginiBlood VesselsRohee TariqÎncă nu există evaluări
- Design and Analysis of A Dual Chamber Cardiac Pacemaker Using VHDL in Biomedical ApplicationDocument3 paginiDesign and Analysis of A Dual Chamber Cardiac Pacemaker Using VHDL in Biomedical ApplicationEditor IJRITCCÎncă nu există evaluări
- QOch 11 Studyguide QODocument16 paginiQOch 11 Studyguide QOHenry HoÎncă nu există evaluări
- Baska MaskDocument8 paginiBaska MaskAnish H DaveÎncă nu există evaluări
- CARDIAC ELECTROPHYSIOLOGY AND THE ELECTROCARDIOGRAM - ClinicalKey PDFDocument40 paginiCARDIAC ELECTROPHYSIOLOGY AND THE ELECTROCARDIOGRAM - ClinicalKey PDFMelanie DascăluÎncă nu există evaluări
- PONS: NeuroanatomyDocument20 paginiPONS: NeuroanatomyHassan IlyasÎncă nu există evaluări
- Important SEQs Heart Physiology by Dr. RoomiDocument3 paginiImportant SEQs Heart Physiology by Dr. RoomiMudassar Roomi100% (2)
- Classification of Nerve FiberDocument21 paginiClassification of Nerve FiberNayab IftikharÎncă nu există evaluări
- Ventricular Assist DeviceDocument12 paginiVentricular Assist DevicesamadonyÎncă nu există evaluări
- U6C6 - Carty Soriano - Heart ValvesDocument6 paginiU6C6 - Carty Soriano - Heart ValvesXio LinaresÎncă nu există evaluări
- Elasticity, Surfactant, Surface Tension and Compliance: Prof - Hafeezul HassanDocument26 paginiElasticity, Surfactant, Surface Tension and Compliance: Prof - Hafeezul HassanDr. Zaheer AliÎncă nu există evaluări
- Lungs & Pleura PDFDocument24 paginiLungs & Pleura PDFEl SpinnerÎncă nu există evaluări
- Pressure Volume LoopDocument4 paginiPressure Volume LoopVinodh PerumalÎncă nu există evaluări
- Cardiac AnatomyDocument6 paginiCardiac Anatomysandranamahen2Încă nu există evaluări
- Lecture 6 Cardiovascular: Vascular System - The HemodynamicsDocument70 paginiLecture 6 Cardiovascular: Vascular System - The HemodynamicsAndreea ŞtefănescuÎncă nu există evaluări
- Contrast Agents, Cath Lab & Anaphylatic Shock-ParamedDocument29 paginiContrast Agents, Cath Lab & Anaphylatic Shock-ParamedManikanta GupthaÎncă nu există evaluări
- Biology: Circulation and Cardiovascular SystemsDocument59 paginiBiology: Circulation and Cardiovascular SystemsIain Choong WKÎncă nu există evaluări
- Fibrous Skeleton of Heart PDFDocument2 paginiFibrous Skeleton of Heart PDFMeghan0% (1)
- ElectrochemistryDocument5 paginiElectrochemistrydanielmahsaÎncă nu există evaluări
- Grip Strength and ElectromyogramDocument8 paginiGrip Strength and ElectromyogramChandra Murali100% (1)
- Composition and Function of Blood ComponentsDocument17 paginiComposition and Function of Blood ComponentsPrakash Kumar Nayak100% (1)
- Frank-Starling LawDocument5 paginiFrank-Starling LawNTA UGC-NETÎncă nu există evaluări
- Memorias FloydDocument36 paginiMemorias FloydDr.R.Udaiya Kumar Professor & Head/ECEÎncă nu există evaluări
- Chapter 11-The Normal ElectrocardiogramDocument18 paginiChapter 11-The Normal Electrocardiogrammuna s100% (1)
- Heart Anatomy: LocationDocument45 paginiHeart Anatomy: LocationZaira100% (1)
- Circulatory System - Part 2 4-8-14 For BBDocument23 paginiCirculatory System - Part 2 4-8-14 For BBroman7dbÎncă nu există evaluări
- Gross Anatomy of Posterior Compartment of The ThighDocument20 paginiGross Anatomy of Posterior Compartment of The ThighOnah ArinzeÎncă nu există evaluări
- Body Plethysmography AvneetDocument54 paginiBody Plethysmography Avneetsalimtajamul100% (3)
- Oxygen CascadeDocument12 paginiOxygen Cascadehy3pjb1984Încă nu există evaluări
- Regulation RespirationDocument34 paginiRegulation Respirationkiedd_04100% (2)
- Blood Vessels Lecture Slides PDFDocument32 paginiBlood Vessels Lecture Slides PDFRaluca AlexandraÎncă nu există evaluări
- Focal Atrial Tachycardia I: Clinical Features, DiagnosisDocument10 paginiFocal Atrial Tachycardia I: Clinical Features, Diagnosisapi-26166949Încă nu există evaluări
- Peripheral Nerve Injury and Repair: Adam Osbourne, 5th Year MedicineDocument5 paginiPeripheral Nerve Injury and Repair: Adam Osbourne, 5th Year Medicineayrahma29Încă nu există evaluări
- Electrocardiogram: What Is An ECG?Document8 paginiElectrocardiogram: What Is An ECG?Ali dhyaa100% (1)
- Cardiology. Section 1. Anatomy of HeartDocument13 paginiCardiology. Section 1. Anatomy of HeartJennifer SteffiÎncă nu există evaluări
- The Circulatory System BookletDocument30 paginiThe Circulatory System BookletTroy CampbellÎncă nu există evaluări
- Nerve StructureDocument53 paginiNerve StructureMedulla Oblongata100% (1)
- Cardiac PacemakersDocument206 paginiCardiac PacemakersMiguel PenarandaÎncă nu există evaluări
- Introduction To Radiology: As Clinical AnatomyDocument30 paginiIntroduction To Radiology: As Clinical AnatomyrizsamaruÎncă nu există evaluări
- Experiment 6:: Electrocardiogram (ECG) and Heart SoundsDocument5 paginiExperiment 6:: Electrocardiogram (ECG) and Heart SoundsWalaa AlzoubiÎncă nu există evaluări
- DR Jagdish Narayan Assistant Professor Department of PhysiologyDocument75 paginiDR Jagdish Narayan Assistant Professor Department of PhysiologyYordanos GetachewÎncă nu există evaluări
- Electronic Spirometer For The Assessment of Lung's FunctionalityDocument4 paginiElectronic Spirometer For The Assessment of Lung's Functionalitynagarajka100% (2)
- Cardiac Muscle: Dr. Kanimozhi Sadasivam, MD Associate Professor SRM Medical College & RC, ChennaiDocument57 paginiCardiac Muscle: Dr. Kanimozhi Sadasivam, MD Associate Professor SRM Medical College & RC, ChennaiShampa SenÎncă nu există evaluări
- Chapter 22 - Structure and Function of The Cardiovascular and Lymphatic Systems - Nursing Test BanksDocument36 paginiChapter 22 - Structure and Function of The Cardiovascular and Lymphatic Systems - Nursing Test BanksNeLÎncă nu există evaluări
- CHAPTER 6 ANAPHY TransesDocument13 paginiCHAPTER 6 ANAPHY TransesHoly HaeinÎncă nu există evaluări
- မိခင္မ်ားအတြက္ ကိုယ္လက္ေလ့က်င့္ခန္းDocument17 paginiမိခင္မ်ားအတြက္ ကိုယ္လက္ေလ့က်င့္ခန္းPan NweÎncă nu există evaluări
- Management of Maxillofacial Trauma in Emergency An Update of Challenges and ControversiesDocument17 paginiManagement of Maxillofacial Trauma in Emergency An Update of Challenges and ControversiesHelna AmeliaÎncă nu există evaluări
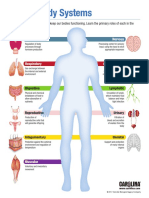
- Human Body System InfographicDocument12 paginiHuman Body System InfographicDiana VillanuevaÎncă nu există evaluări
- Design and Fabrication of A Full Denture With Shorthened Dental Arch and Flat Ridge in MandibularDocument10 paginiDesign and Fabrication of A Full Denture With Shorthened Dental Arch and Flat Ridge in MandibulardinaÎncă nu există evaluări
- ArnoldDocument3 paginiArnoldGiambattista Mastrapasqua100% (1)
- Endocrinotox Vademecum: Homeopathic Regulation of The Neuroendocrine SystemDocument77 paginiEndocrinotox Vademecum: Homeopathic Regulation of The Neuroendocrine SystemlaviniagavrilaÎncă nu există evaluări
- Workout PlannerDocument9 paginiWorkout PlannerAi JoieÎncă nu există evaluări
- Topic 2.1 Ventilatory System Sport Science IBDocument4 paginiTopic 2.1 Ventilatory System Sport Science IBAntonio MouÎncă nu există evaluări
- Skeletal System (Sem 1)Document231 paginiSkeletal System (Sem 1)joel david knda mjÎncă nu există evaluări
- June 17 26 s001 PDFDocument9 paginiJune 17 26 s001 PDFBetul RojeabÎncă nu există evaluări
- SAUDI Licensing Orthodontics ExaminationDocument34 paginiSAUDI Licensing Orthodontics ExaminationRamy Fathy100% (1)
- 8.anterior GuidanceDocument15 pagini8.anterior GuidanceVikas AggarwalÎncă nu există evaluări
- Asda JKLMNDocument185 paginiAsda JKLMNBikram SinghÎncă nu există evaluări
- 2021 02 Anatomy OutlineDocument5 pagini2021 02 Anatomy OutlineCinja ShidoujiÎncă nu există evaluări
- Histology of The EyeDocument91 paginiHistology of The EyeFazira EkmaÎncă nu există evaluări
- Traumatic Tympanic Membrane PerforationDocument7 paginiTraumatic Tympanic Membrane PerforationNada ZultiÎncă nu există evaluări
- A Prospective Study in Analgesic Effect of Distal Sodium Channel Blockers (DSCB) in Patients With SciaticaDocument5 paginiA Prospective Study in Analgesic Effect of Distal Sodium Channel Blockers (DSCB) in Patients With SciaticaInternational Journal of Innovative Science and Research Technology100% (1)
- Neuroscience - 4.3 - Examination of Cerebellar Systems and Meninges (KSD)Document4 paginiNeuroscience - 4.3 - Examination of Cerebellar Systems and Meninges (KSD)Kevin C. AguilarÎncă nu există evaluări
- Maglaqui - Skeletal SystemDocument5 paginiMaglaqui - Skeletal SystemNICOLE MAGLAQUIÎncă nu există evaluări
- Vedic Healing Through Gems4Document3 paginiVedic Healing Through Gems4gesÎncă nu există evaluări
- TNCC NotesDocument9 paginiTNCC NotesJessie Bronkalla95% (21)
- Development of The Digestive SystemDocument90 paginiDevelopment of The Digestive Systemyusrah mukhtarÎncă nu există evaluări
- Body Beast Workout SheetsDocument24 paginiBody Beast Workout Sheetssaf09409tÎncă nu există evaluări
- 9) Cardio Vascular AssessmentDocument40 pagini9) Cardio Vascular Assessmentrajaguru2000393% (15)
- Neuroanatomy An Illustrated Colour Text Crossman 6 Ed 2020 PDFDocument185 paginiNeuroanatomy An Illustrated Colour Text Crossman 6 Ed 2020 PDFKatrin CoolChe100% (3)
- Anatomy Muscles of The FaceDocument7 paginiAnatomy Muscles of The Facecristienne marielle MercadoÎncă nu există evaluări
- Diagnosis TopisDocument101 paginiDiagnosis TopisGita Nur SiwiÎncă nu există evaluări
- Ornac Standards PositioningDocument7 paginiOrnac Standards PositioningAndika X NandaÎncă nu există evaluări