S-ar putea să vă placă și
- The Placebo Effect in Popular Culture: Mary Faith MarshallDocument6 paginiThe Placebo Effect in Popular Culture: Mary Faith MarshallRajan PandaÎncă nu există evaluări
- s0308 8146 2801 2900391 0Document5 paginis0308 8146 2801 2900391 0Rajan PandaÎncă nu există evaluări
- Mask Maker, Mask Maker: The Black Gay Subject in 1970S Popular CultureDocument30 paginiMask Maker, Mask Maker: The Black Gay Subject in 1970S Popular CultureRajan PandaÎncă nu există evaluări
- Popular Culture Representations of Science: Views From The Canadian Stem Cell Research CommunityDocument3 paginiPopular Culture Representations of Science: Views From The Canadian Stem Cell Research CommunityRajan PandaÎncă nu există evaluări
- s0300 483x 2802 2900751 5Document9 paginis0300 483x 2802 2900751 5Rajan PandaÎncă nu există evaluări
- Can QOL Researchers Learn Anything Scientifically'' Meaningful From Popular Cultural Icons Who Speak On Happiness and Well-Being?Document3 paginiCan QOL Researchers Learn Anything Scientifically'' Meaningful From Popular Cultural Icons Who Speak On Happiness and Well-Being?Rajan PandaÎncă nu există evaluări
- Modernity East and West: Melodrama And: Yanqing in Shanghai's Popular CultureDocument20 paginiModernity East and West: Melodrama And: Yanqing in Shanghai's Popular CultureRajan PandaÎncă nu există evaluări
- Biology in The Movies: Using The Double-Edged Sword of Popular Culture To Enhance Public Understanding of ScienceDocument6 paginiBiology in The Movies: Using The Double-Edged Sword of Popular Culture To Enhance Public Understanding of ScienceRajan PandaÎncă nu există evaluări
- The Symposium On Urban Popular Culture in Modern China: MA Min, JIANG Jin, WANG Di, Joseph W. ESHERICK, LU HanchaoDocument34 paginiThe Symposium On Urban Popular Culture in Modern China: MA Min, JIANG Jin, WANG Di, Joseph W. ESHERICK, LU HanchaoRajan PandaÎncă nu există evaluări
- Book Reviews: Beyond Sexuality. by Tim Dean. University of ChicagoDocument9 paginiBook Reviews: Beyond Sexuality. by Tim Dean. University of ChicagoRajan PandaÎncă nu există evaluări
- Burnout: From Popular Culture To Psychiatric Diagnosis in SwedenDocument21 paginiBurnout: From Popular Culture To Psychiatric Diagnosis in SwedenRajan PandaÎncă nu există evaluări
- Profile of Nonprotein Thiols, Lipid Peroxidation and D-Aminolevulinate Dehydratase Activity in Mouse Kidney and Liver in Response To Acute Exposure To Mercuric Chloride and Sodium SeleniteDocument9 paginiProfile of Nonprotein Thiols, Lipid Peroxidation and D-Aminolevulinate Dehydratase Activity in Mouse Kidney and Liver in Response To Acute Exposure To Mercuric Chloride and Sodium SeleniteRajan PandaÎncă nu există evaluări
- Culture and Stigma: Popular Culture and The Case of Comic BooksDocument28 paginiCulture and Stigma: Popular Culture and The Case of Comic BooksRajan PandaÎncă nu există evaluări
- s11002 005 2774 6Document12 paginis11002 005 2774 6Rajan PandaÎncă nu există evaluări
- s0304 422x 2803 2900036 6Document18 paginis0304 422x 2803 2900036 6Rajan PandaÎncă nu există evaluări
- s0308 8146 2803 2900030 XDocument5 paginis0308 8146 2803 2900030 XRajan PandaÎncă nu există evaluări
- Modulation of Collagen-Induced Arthritis by IL-4 and Dexamethasone: The Synergistic Effect of IL-4 and Dexamethasone On The Resolution of CIADocument8 paginiModulation of Collagen-Induced Arthritis by IL-4 and Dexamethasone: The Synergistic Effect of IL-4 and Dexamethasone On The Resolution of CIARajan PandaÎncă nu există evaluări
- Drying Kinetics of Fresh and Osmotically Pre-Treated Papaya (Carica Papaya L.)Document7 paginiDrying Kinetics of Fresh and Osmotically Pre-Treated Papaya (Carica Papaya L.)Rajan PandaÎncă nu există evaluări
- s0167 4048 2800 2904010 4Document2 paginis0167 4048 2800 2904010 4Rajan PandaÎncă nu există evaluări
- s0145 2126 2803 2900013 4Document7 paginis0145 2126 2803 2900013 4Rajan PandaÎncă nu există evaluări
- s0308 8146 2800 2900134 5Document5 paginis0308 8146 2800 2900134 5Rajan PandaÎncă nu există evaluări
- US DoD Cybercrime Database to Aid Security EffortsDocument3 paginiUS DoD Cybercrime Database to Aid Security EffortsRajan PandaÎncă nu există evaluări
- s0097 8485 2802 2900029 3Document32 paginis0097 8485 2802 2900029 3Rajan PandaÎncă nu există evaluări
- s0040 4039 2801 2996980 1Document4 paginis0040 4039 2801 2996980 1Rajan PandaÎncă nu există evaluări
- s0031 9422 2899 2900177 6Document5 paginis0031 9422 2899 2900177 6Rajan PandaÎncă nu există evaluări
- s0031 9422 2800 2900295 8Document4 paginis0031 9422 2800 2900295 8Rajan PandaÎncă nu există evaluări
- E Ects of Lignoids On A Hematophagous Bug, Rhodnius Prolixus: Feeding, Ecdysis and DiuresisDocument6 paginiE Ects of Lignoids On A Hematophagous Bug, Rhodnius Prolixus: Feeding, Ecdysis and DiuresisRajan PandaÎncă nu există evaluări
- Electrochemical Aspects of The Reduction of Biologically Active 2-Hydroxy-3-Alkyl-1,4-NaphthoquinonesDocument12 paginiElectrochemical Aspects of The Reduction of Biologically Active 2-Hydroxy-3-Alkyl-1,4-NaphthoquinonesRajan PandaÎncă nu există evaluări
- Ion Exchange Between Divalent Counterions in Anionic Micellar SolutionDocument5 paginiIon Exchange Between Divalent Counterions in Anionic Micellar SolutionRajan PandaÎncă nu există evaluări
- Shoe Dog: A Memoir by the Creator of NikeDe la EverandShoe Dog: A Memoir by the Creator of NikeEvaluare: 4.5 din 5 stele4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDe la EverandGrit: The Power of Passion and PerseveranceEvaluare: 4 din 5 stele4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDe la EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceEvaluare: 4 din 5 stele4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)De la EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Evaluare: 4 din 5 stele4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDe la EverandThe Little Book of Hygge: Danish Secrets to Happy LivingEvaluare: 3.5 din 5 stele3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealDe la EverandOn Fire: The (Burning) Case for a Green New DealEvaluare: 4 din 5 stele4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDe la EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeEvaluare: 4 din 5 stele4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDe la EverandNever Split the Difference: Negotiating As If Your Life Depended On ItEvaluare: 4.5 din 5 stele4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDe la EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureEvaluare: 4.5 din 5 stele4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDe la EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryEvaluare: 3.5 din 5 stele3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDe la EverandThe Emperor of All Maladies: A Biography of CancerEvaluare: 4.5 din 5 stele4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDe la EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreEvaluare: 4 din 5 stele4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDe la EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyEvaluare: 3.5 din 5 stele3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDe la EverandTeam of Rivals: The Political Genius of Abraham LincolnEvaluare: 4.5 din 5 stele4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDe la EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersEvaluare: 4.5 din 5 stele4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDe la EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaEvaluare: 4.5 din 5 stele4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaDe la EverandThe Unwinding: An Inner History of the New AmericaEvaluare: 4 din 5 stele4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)De la EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Evaluare: 4.5 din 5 stele4.5/5 (119)
- Her Body and Other Parties: StoriesDe la EverandHer Body and Other Parties: StoriesEvaluare: 4 din 5 stele4/5 (821)
- Function: What Is The Skeletal System?Document6 paginiFunction: What Is The Skeletal System?Mr. Christian ParabuacÎncă nu există evaluări
- Google CardboardDocument3 paginiGoogle CardboardMartínJiménezÎncă nu există evaluări
- The Differential Diagnosis of Fluoride and Non-Fluoride OpacitiesDocument4 paginiThe Differential Diagnosis of Fluoride and Non-Fluoride OpacitiesRajshekhar BanerjeeÎncă nu există evaluări
- Congenital LaryngomalaciaDocument8 paginiCongenital LaryngomalaciaRettha SigiroÎncă nu există evaluări
- AIIMS Bibinagar Recruitment for Faculty PostsDocument22 paginiAIIMS Bibinagar Recruitment for Faculty PostsavinashÎncă nu există evaluări
- PORNOGRAPHICDocument13 paginiPORNOGRAPHICcarlos ortizÎncă nu există evaluări
- MsdsDocument6 paginiMsdsJackyÎncă nu există evaluări
- Digestive System PowerpointDocument33 paginiDigestive System PowerpointThomas41767% (6)
- Bmjopen 2017 016402Document6 paginiBmjopen 2017 016402Ćatke TkećaÎncă nu există evaluări
- Understanding Uterine FibroidsDocument52 paginiUnderstanding Uterine FibroidsDoctor JitÎncă nu există evaluări
- Adult Health - Soap Note 5Document3 paginiAdult Health - Soap Note 5api-546259691100% (3)
- 11 Foods That Lower Cholesterol: Foods That Make Up A Low Cholesterol Diet Can Help Reduce High LevelsDocument3 pagini11 Foods That Lower Cholesterol: Foods That Make Up A Low Cholesterol Diet Can Help Reduce High Levelsaj dancel marcosÎncă nu există evaluări
- OPD Network ListDocument354 paginiOPD Network ListSHAIKH ABDUL AZIZ salim bashaÎncă nu există evaluări
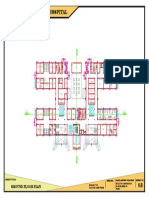
- 100-Bed General Hospital LayoutDocument1 pagină100-Bed General Hospital LayoutAshish chauhanÎncă nu există evaluări
- Ergonomics (Uu Tien Dich)Document6 paginiErgonomics (Uu Tien Dich)Phong D LeÎncă nu există evaluări
- KPI - Foruth EditionDocument30 paginiKPI - Foruth EditionAnonymous qUra8Vr0SÎncă nu există evaluări
- DSM 5Document35 paginiDSM 5Hemant KumarÎncă nu există evaluări
- ASP ProductsDocument33 paginiASP ProductsSerghei Barba100% (1)
- Crodua Prioritization TableDocument10 paginiCrodua Prioritization TableThea DuoÎncă nu există evaluări
- Complete VaccinationDocument2 paginiComplete VaccinationNgoo NwosuÎncă nu există evaluări
- Stress Amongst Healthcare Professionals and Migrant Workers During Covid-19 PandemicDocument6 paginiStress Amongst Healthcare Professionals and Migrant Workers During Covid-19 PandemicIJAR JOURNALÎncă nu există evaluări
- Introduction To Public Health... 1stDocument37 paginiIntroduction To Public Health... 1stNELSONJD20195100% (3)
- GENETIC DISORDERS AND CYTOGENETICSDocument7 paginiGENETIC DISORDERS AND CYTOGENETICSsmilechance8Încă nu există evaluări
- S.P.P.M. MasterDocument126 paginiS.P.P.M. MasterwahyuÎncă nu există evaluări
- How To Make An ELearning ModuleDocument22 paginiHow To Make An ELearning ModulePradeep RawatÎncă nu există evaluări
- Methodological Literature Review 1 1Document8 paginiMethodological Literature Review 1 1api-584018105Încă nu există evaluări
- Domestic Physician HeringDocument490 paginiDomestic Physician Heringskyclad_21Încă nu există evaluări
- Hahnemann Advance MethodDocument2 paginiHahnemann Advance MethodRehan AnisÎncă nu există evaluări
- Artikel 3Document23 paginiArtikel 3Hadian UwuoÎncă nu există evaluări
- 10 Doh Approved Herbal MedicineDocument32 pagini10 Doh Approved Herbal MedicineIsmael JaaniÎncă nu există evaluări